El árbol de la vida: el Cerebelo
Si se secciona el cerebelo por la línea media se puede observar que la mayor parte de la sustancia gris se sitúa en la superficie formando una estructura, denominada corteza cerebelosa, que envuelve la sustancia blanca. Ésta se ramifica hacia la sustancia gris y aporta al cerebelo la forma arbórea que hace que sea llamado “el árbol de la vida”.
La corteza del cerebelo es una estructura laminada formada por tres capas horizontales en cualquier zona de la superficie del cerebelo que se observe.
La capa más interna es la capa granular en la que hay un gran número de interneuronas: las células granulares y las células de Golgi. Hay tantas células granulares en el cerebelo que se estima que su número sobrepasa el de todas las células de la corteza cerebral. Los axones de las granulares son muy finos y se denominan fibras paralelas porque después de ascender a la capa más superficial se dividen en dos ramas paralelas a los pliegues. Esta capa superficial se denomina capa molecular y sus células características son también interneuronas: las células estrelladas y las células en cesto.
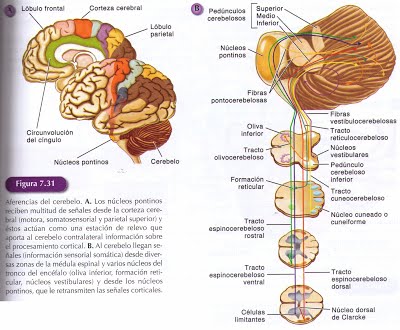 La capa de células de Purkinge se localiza entre las dos anteriores y está formada por los somas de las células de Purkinge que son muy grandes y numerosos y se disponen en una única fila dentro de la capa.
La capa de células de Purkinge se localiza entre las dos anteriores y está formada por los somas de las células de Purkinge que son muy grandes y numerosos y se disponen en una única fila dentro de la capa.
Forman filas perpendiculares a las fibras paralelas, con loas que establecen sinápsis sus dendritas, y a través de ellas reciben muchas de las señales que llegan a la corteza cerebelosa desde la médula espinal, la formación reticular y los núcleos pontinos del tronco del encéfalo. Las células de Purkinge son las únicas células de proyección de la corteza cerebelosa, y sus axones se dirigen, formando parte de la sustancia blanca, hasta los núcleos profundos del cerebelo.
Por los núcleos profundos del cerebelo pasan todas las señales que salen de la corteza cerebelosa y también muchas de las que llegan a ella. Estos núcleos constituyen una pequeña parte de la sustancia gris del cerebelo y están inmersos en la sustancia blanca. Se localizan próximos al techo del IV ventrículo muy distantes de la corteza cerebelosa, a cada lado de un eje central imaginario, el más cercano a la línea media es el núcleo fastígeo, próximos a éste se localizan los núcleos interpuestos y lateral a ellos se localiza el núcleo dentado.
Esta situación medio-lateral de los núcleos profundos es equivalente a la de las tres zonas longitudinales de la corteza del cerebelo y cada uno se relaciona con la zona de localización equivalente de la corteza. Se forman así tres zonas longitudinales en el cerebelo desde la corteza a los núcleos profundos y viceversa. Desde cada una de las tres zonas de la corteza cerebelosa, los axones de las células de Purkinje convergen sobre sus correspondientes núcleos profundos y en éstos se originan las señales que salen de las distintas zonas del cerebelo, formando así, distintas unidades funcionales. Desde la zona lateral del cerebelo (zona lateral de los hemisferios-núcleo dentado) las señales se dirigen, a través del tálamo, a la corteza cerebral y como a su vez, recibe muchas señales corticales a través de los núcleos pontinos, por esta interrelación con la corteza cerebral, esta zona se denomina cerebrocerebelo. Las señales de las zonas intermedia y medial del cerebro (zona intermedia de los hemisferios -núcleos interpuestos y vermis- núcleo fastigio) terminan en varios núcleos del tronco del encéfalo, que envían proyecciones que descienden a la médula espinal, y reciben muchas señales desde ésta, por lo que se denominan espinocerebelo. Los axones del lóbulo floculonodular se dirigen directamente a los núcleos vestibulares ipsilaterales, de los que reciben numerosas señales, por lo que se denomina vestibulocerebelo. Tanto en las áreas corticales, como en estos núcleos del tronco del encéfalo, que reciben señales desde el cerebelo, se originan tractos descendentes que intervienen en el control motor y su actividad está modulada por estas tres unidades funcionales del cerebelo.
La nuez más seleccionada: organización interna de los hemisferios cerebrales
Al igual que en el cerebelo, la superficie externa de los hemisferios cerebrales está formada por el manto de sustancia gris que forma la corteza cerebral. Por debajo de la corteza cerebral se encuentra la sustancia blanca y embebidas en la profundidad de la sustancia blanca están las estructuras subcorticales.
Los hemisferios cerebrales y el cerebelo son similares en estos aspectos, pero su organización anatómica y funcional es mucho más compleja que la del cerebelo porque tiene muchas más estructuras subcorticales y porque la corteza cerebral tiene más capas, y éstas tienen características propias en las diferentes zonas.
 Las estructuras subcorticales
Las estructuras subcorticales
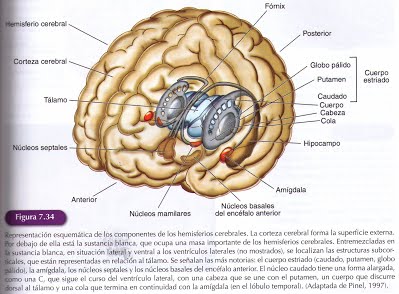
En secciones coronales de los hemisferios cerebrales las estructuras subcorticales se observan inmersas en la sustancia blanca, organizadas alrededor de los ventrículos laterales y bordeando el diencéfalo.
En el centro de los hemisferios cerebrales, lateral al ventrículo lateral, está el cuerpo estriado que agrupa tres grandes núcleos subcorticales: el núcleo caudado, el putamen y el globo pálido. El putamen es el mayor de los núcleos subcorticales y se localiza lateralmente bajo el lóbulo de la ínsula. Está separado del núcleo caudado por la cápsula interna. los núcleos caudado y putamen forman el neoestriado. El tercer componente del cuerpo estriado es el globo pálido que se localiza adyacente al putamen. Unas delgadas láminas lo parcelan en dos segmentos: lateral y medial, teniendo cada segmento su propia conectividad.
El cuerpo estriado tiene una gran relación anatomo-funcional con la sustancia negra (mesencéfalo) y el núcleo subtalámico (diencéfalo) y juntos se denominan con el término de ganglios basales. En la zona ventral del ventrículo lateral se localiza otra estructura subcortical, el núcleo de la estría terminal que está atravesado por la comisura anterior y queda separado lateralmente del globo pálido por la capsula interna. En la cara medial del ventrículo lateral están los núcleos septales, que quedan adyacentes al fórnix. En la parte ventral de los hemisferios, bordeando estructuras diencefálicas, hay varios núcleos que, en su conjunto, se refieren como núcleos basales del encéfalo anterior. Entre ellos, está el núcleo basal de Meynert que tiene proyecciones que se distribuyen ampliamente por la corteza cerebral y la degeneración de sus neuronas está relacionada con la enfermedad de Alzheimer.
En la profundidad del lóbulo tempora, situada debajo de la corteza olfatoria se encuentra la amígdala, que está formada por varios núcleos que se agrupan en tres unidades funcionales diferentes. La amígdala, los núcleos septales y el núcleo de la estría terminal forman parte de circuitos neurales que controlan conductas emocionales y motivadas, como el miedo, la agresividad o la conducta sexual.
La sustancia blanca
La sustancia blanca ocupa gran parte de los hemisferios cerebrales. Está formada por una inmensa cantidad de axones que, en su mayoría, se agrupan en grandes tractos que son visibles a simple vista cuando se observan los hemisferios cerebrales. Entre los diversos componentes de los hemisferios cerebrales se establece una conectividad muy compleja. Hay numerosas vía de comunicación entre la corteza cerebral y las estructuras subcorticales, y otras tantas que se circunscriben a nivel subcortical. Los hemisferios cerebrales mantienen también una gran comunicación con las divisiones más caudales del encéfalo y con la médula espinal. En relación con su función los tractos (fibras) de los hemisferios cerebrales son de tres tipos.
- Las comisuras interhemisféricas son las que están formadas por axones que interconectan estructuras de ambos hemisferios. Las principales son la comisura anterior que interconecta los bulbos olfatorios y las regiones inferiores del lóbulo temporal de ambos hemisferios cerebrales, y el cuerpo calloso. El cuerpo calloso es la comisura más grande y une ambos hemisferios por debajo de la cisura longitudinal y por encima de los ventrículos laterales. Se estima que los axones que lo componen pueden alcanzar los 300 millones, y están mielinizados, por lo que adquiere una apariencia blanquecina y compacta. El cuerpo calloso se abre radialmente hacia los laterales de los hemisferios. Muchas de sus fibras interconectan regiones de la corteza cerebral localizadas en el mismo lugar de cada lóbulo de ambos hemisferios y cuando se secciona el cuerpo calloso se impide la transferencia de información de un hemisferio a otro.
- Los tractos o fibras de proyección. Los mayores son los que agrupan los axones que ascienden hasta la corteza cerebral y los que ésta envía a las regiones más caudales del encéfalo y la médula espinal. Estos tractos son: la corona radiada, que está formada por axones que se distribuyen por la parte superior de los hemisferios cerebrales en forma de abanico, y la capsula interna que recoge ya los axones en un tracto más compacto que se adentra en los pedúnculos cerebrales al llegar al mesencéfalo. Otra tracto de proyección importante es el fórnix, que conecta la formación hipocampal con núcleos subcorticales (los núcleos septales) y con el hipotálamo (los núcleos mamilares)
- Las fibras de asociación que son los axones que conectan distintas zonas de la corteza cerebral del mismo hemisferio. En cada hemisferio cerebral existen muchos haces de axones que interconectan diversas regiones de la corteza cerebral del mismo lado. Los que conectan regiones de circunvoluciones adyacentes se denominan fibras de asociación cortas o fibras U, porque al arquearse bajo las cisuras toman la forma de esta vocal. Pero también existe una profusa comunicación entre regiones mucho más distantes, a estas fibras se las denomina fibras de asociación largas.
La corteza cerebral
La corteza cerebral es una estructura que forma la superficie externa de los hemisferios cerebrales.
Tiene las células organizadas en capas horizontales a la superficie de los hemisferios. Sin embargo, tanto las células características como los circuitos que se establecen en su interior, son muy distintas a las células de Purkinge y los circuitos del cerebelo. Las células de proyección típicas de la corteza cerebral son las células piramidales, que son características de la corteza cerebral de todos los vertebrados. No todas las zonas de la corteza cerebral tienen la misma organización, ya que hay variabilidad de capas entre las diversas zonas de la misma. Podemos distinguir dos tipos de corteza cerebral: la allocorteza y la neocorteza.
La Allocorteza
La corteza filogenéticamente más antigua es la corteza olfatoria (paleocorteza) y la corteza del lóbulo límbico (arquicorteza). Se denomina globalmente allocorteza y es el tipo de corteza dominante en los vertebrados inferiores, en los humanos representa aproximadamente el 10%. Se organiza en un número variable de capas. En la cara medial del lóbulo temporal está organizada en tres capas. Esta zona de la corteza pertenece a la formación hipocampal que, en cortes coronales tiene una forma característica similar a un “brazo de gitano” ya que la corteza se pliega sobre sí misma.
En la formación hipocampal se incluyen el hipocampo, el giro dentado, el complejo del subículo y la corteza entorrinal. Al hipocampo también se le denomina cuerno o asta de Ammon porque en continuidad con el fórnix que es el principal tracto de proyección de la formación hipocampal, se parece a las astas de un macho cabrío. Es un componente fundamental de los circuitos neurales que intervienen en los procesos de aprendizaje y la memoria, entre ellos de la memoria espacial.
La Neocorteza
La neocorteza constituye aproximadamente el 90% de la corteza cerebral humana, por lo que forma la superficie externa de la mayor parte de los hemisferios cerebrales. Se denomina corteza nueva porque tuvo un gran desarrollo en el evolución reciente, con la aparición de los mamíferos, lo que propició un aumento importante en el tamaño de los hemisferios cerebrales. Esta corteza llega a representar más de la mitad de toda la sustancia gris del SNC. Dado que supone un gran porcentaje de la corteza cerebral, habitualmente se le denominará corteza cerebral. Está organizada en seis capas. Sus células de proyección características, las células piramidales tienen dendritas apicales que están en contacto con la capa superficial, y su axón se extiende por otras capas o pasa a formar parte de la sustancia blanca. En las seis capas que la componen hay otros tipos de células que son fundamentalmente interneuronas. Las seis capas se diferencian por su tipo celular característico y por la densidad.
Las capas I, II, IV son fundamentalmente, capas receptoras de las aferencias corticales y cada una recibe aferencias originadas en distintas zonas del SNC, es decir, que se produce una separación de las aferencias por capas. Las aferencias de la corteza se originan principalmente en las capas II, III, V y VI, aunque la capa IV es fundamentalmente receptora, también es efectora.
Características fundamentales de la organización cortical
Una característica fundamental de la organización cortical es la interacción horizontal y vertical que se produce entre sus neuronas. Los colaterales axónicos de las neuronas piramidales y los axones de las interneuronas, que se disponen en distintas orientaciones, permiten que se produzca una gran interacción vertical y horizontal entre las neuronas corticales. De este modo, a pesar de que las aferencias corticales se segreguen por capas, las redes densas y enmarañadas que forman las prolongaciones de los axones y las dendritas (neuropilo) de la corteza cerebral permiten que la activación se propague en zonas corticales adyacentes. Estas interacciones entre las células corticales constituyen un aspecto importante de la organización antomofuncional de la corteza cerebral, ya que son la base anatómica de los procesos fisiológicos fundamentales.
Otra característica es que sus células tienen una marcada tendencia a organizarse verticalmente. Es lo que se denomina organización columnar, se forman pequeños cilindros o columnas de tejido cortical que atraviesan el espesor de las capas de la corteza a modo de radios perpendiculares a la superficie. Las neuronas forman columnas de dimensiones micrométricas, que tienen propiedades funcionales distintas de las columnas adyacentes. Esta organización columnar la descubrió Mountcastle.
Hay que añadir también, que la neocorteza no es uniforme, existe diferencia en el grosor y la estructura de las capas entre las diferentes zonas. Estas diferencias estructurales se han utilizado para realizar numerosos mapas citoarquitectónicos que parcelan la corteza cerebral en áreas con características estructurales distintas.
Áreas de la Neocorteza
El mapa citoarquitectónico más utilizado es el realizado por Brodmann, dividió la corteza cerebral en más de 50 áreas con diferencias estructurales, y sugirió que las diferentes áreas realizaban también diferentes funciones. Actualmente está comprobado que diferentes áreas de la corteza tienen diferencias estructurales y realizan funciones distintas.
Las áreas sensoriales de la corteza cerebral llevan a cabo el procesamiento superior de la información de las distintas modalidades sensoriales, de modo que cada sentido tiene sus propias áreas sensoriales. Hay áreas sensoriales primarias y secundarias. Las áreas sensoriales primarias son las que reciben la mayoría de sus aferencias sensoriales directamente desde los núcleos de relevo sensorial del tálamo y cada sentido tiene su área sensorial primaria. Próximas a ellas, están las áreas sensoriales secundarias o áreas sensoriales de orden superior, que son las que reciben la mayoría de sus aferencias desde sus correspondientes áreas sensoriales primarias o desde otras áreas sensoriales secundarias del mismo sentido, y se ocupan de aspectos elaborados del procesamiento de la información sensorial.
En el lóbulo frontal, adyacentes a la cisura central, hay áreas corticales que tienen características citoarquitectónicas diferentes a las anteriores. Las áreas con estas características son áreas motoras, y se clasifican en área motora primaria y áreas premotoras o corteza motora secundaria. El área motora primaria elabora órdenes que envía a través de las vías motoras descendentes hasta las motoneuronas para que éstas ejecuten los movimientos de las diferentes partes del cuerpo.
Las áreas premotoras también originan proyecciones motoras descendentes y, además intervienen en la planificación de los movimientos y envían numerosas proyecciones al área motora primaria.
La mayor parte de la neocorteza no se ajusta ni en su citoarquitectura ni en sus funciones, a esta descripción porque las áreas sensorial y motora representan un porcentaje moderado de la neocorteza. Una gran parte de la neocorteza tiene muy pocas células en las capas IV-V y una gran densidad celular en las capas II y III que son, fundamentalmente las capas que reciben y envían axones a otras zonas de la corteza.
Esta parte de la corteza se denomina corteza de asociación. Mientras que las áreas sensoriales y motoras se ocupan, respectivamente, de codificar la información sensorial y de realizar el control motor, las áreas de asociación realizan funciones de integración.