El supuesto fundamental en que se basa la Psicobiología es que el comportamiento que observamos y los procesos mentales que intervienen decisivamente en la aparición del comportamiento son fruto del funcionamiento del sistema nervioso.
Las características estructurales y funcionales de nuestro SN son el resultado de la evolución. La evolución ha dotado a los organismos de un sistema nervioso que les permite integrar la información que reciben y emitir la respuesta más adecuada para su supervivencia y reproducción. Aunque el desarrollo filogenético dota a todos los miembros de una especie de un patrón común de organización del SN existen diferencias individuales en la mayoría de las conductas, para comprender esas diferencias es necesario considerar otra serie de factores que determinan el funcionamiento del sistema nervioso. La dotación genética es un condicionante biológico a considerar en la explicación del comportamiento ya que en ella quedan plasmados los logros adaptativos de una especie y es una importante fuente de variabilidad entre sus miembros. Aunque el SN se desarrolla según la información contenida en los gens de cada individuo, es necesario tener en cuenta los factores epigenéticos, que procedentes del ambiente externo (estimulación sensorial...) como interno (hormonas...) del individuo en desarrollo, regulan la forma en que la dotación genética se expresa.
Nuestro sistema nervioso está formado por miles de millones de neuronas y otras células auxiliares.
Este sistema complejo se estructura en un diseño complicado pero perfectamente organizado que permite realizar funciones complejas como son captar la información procedente del medio, analizarla, almacenarla, integrarla y organizar una respuesta adecuada para hacer frente a las situaciones cambiantes de la vida.
Nuestro encéfalo se organiza en una jerarquía constituida por diferentes centros y circuitos nerviosos responsables de funciones específicas. El encéfalo junto con la médula espinal forma el sistema nervioso central (SNC). El SNC está en constante comunicación con el resto de nuestro cuerpo a través de vías que le permiten recibir información del medio ambiente interno y externo, ejecutar respuestas motoras y coordinar los demás órganos para mantener las funciones vitales como la respiración y la digestión. Todas estas vías nerviosas externas al encéfalo y la médula espinal forman el sistema nervioso periférico (SNP).
Las protagonistas fundamentales del SN son las neuronas. Conocer la estructura de la neuronas nos ayuda a comprender cómo funcionan individualmente y la forma en que se agrupan en circuitos que median las funciones exclusivas del SN como son, entre otras, el procesamiento de información y destrezas, el habla, el pensamiento, las emociones y la conciencia de la propia existencia.
La neurona
 Las neuronas son los componentes fundamentales y las unidades básicas de procesamiento del SN.
Las neuronas son los componentes fundamentales y las unidades básicas de procesamiento del SN.
El funcionamiento del SN, y en consecuencia la conducta depende de la comunicación que se establece entre neuronas constituyendo complejísimos circuitos neuronales. El encéfalo humano está compuesto aproximadamente por unos cien mil millones de neuronas y cada neurona conecta con otras diez mil neuronas. La función principal de cada neurona consiste en recibir información y transmitirla una vez procesada. La función integradora que realiza cada neurona constituye la base del funcionamiento neuronal.
Las neuronas poseen la misma información genética, tienen los mismos elementos estructurales y realizan las mismas funciones básicas que cualquier otra célula del cuerpo. Para llevar a cabo una función especializada como es el procesamiento de la información, poseen características que las distinguen de otras células: cuentan con una membrana externa que posibilita la conducción de impulsos nerviosos y tiene la capacidad de transmitir la información tanto de una neurona a otra como a otras células de nuestro organismo. Esta transmisión de información recibe el nombre de transmisión sináptica.
El conocimiento de la estructura del tejido nervioso es relativamente reciente. Fue Camilo Golgi quien desarrolló un método de tinción que hacía posible analizar con extraordinaria precisión el tejido nervioso. Este tinte le permitió observar que la célula nerviosa tiene una larga prolongación cilíndrica -el axón- y otras prolongaciones más pequeñas- las dendritas- ; sin embargo no pudo explicar cuál era la función de estas prolongaciones en la relación que se establecía entre células. Mantuvo la teoría de que las neuronas formaban una red a través de la cual se comunicaban por continuidad de forma aleatoria.
Posteriormente Santiago Ramón y Cajal puso de manifiesto que cada célula nerviosa dispone de un campo receptivo (dendritas) un segmento conductor (axón)y un extremo transmisor (terminal axónico) que la neurona es una entidad discreta y bien definida y no una parte de una red continua como proponía Golgi.
Estableció que estas las células separadas se comunicaban entre sí a través de la sinapsis y propuso los principios básicos de la comunicación neuronal:
Principio de polarización dinámica. La comunicación entre neuronas se establece en una dirección, desde el axón de una neurona a las dendritas o soma neuronal de otra.
Principio de especificidad de las conexiones. No hay una continuidad citoplasmática entre las neuronas ya que incluso en el lugar donde se establece la comunicación existe una separación (hendidura sináptica). Esta comunicación se establece de forma altamente organizada de tal manera que cada célula se comunica con células concretas en puntos especializados de contacto sináptico.
La amplia labor investigadora llevada a cabo por Ramón y Cajal aporto la base experimental y conceptual para establecer la Teoría neuronal por la que junto a Golgi obtuvo el Premio Nobel en 1906.
 Nuestro Sistema Nervioso mantiene durante toda la vida del organismo la capacidad de experimentar modificaciones en su organización anatómica y funcional. A esto hace referencia la plasticidad neuronal, a la capacidad de nuestro SN para cambiar y reorganizarse en función de las situaciones que se van produciendo a lo largo de la vida. El mismo Ramón y Cajal planteó la hipótesis de que los cambios observados en el comportamiento humano, tendrían probablemente un sustrato anatómico, hipótesis que en la actualidad se ha confirmado. Estos cambios son mayores en las primeras etapas de nuestra vida durante el desarrollo de circuitos neurales, pero en la madurez se siguen modificando nuestros contactos sinápticos a medida que aprendemos y establecemos memorias nuevas, que nos enfrentamos a situaciones fisiológicas diversas o cuando nuestro SN se ve sometido a enfermedades o lesiones. La capacidad de cambio continuo contribuye que el desarrollo y la organización del SN no esté marcado únicamente por la acción de los genes y que el cerebro de cada uno sea único.
Nuestro Sistema Nervioso mantiene durante toda la vida del organismo la capacidad de experimentar modificaciones en su organización anatómica y funcional. A esto hace referencia la plasticidad neuronal, a la capacidad de nuestro SN para cambiar y reorganizarse en función de las situaciones que se van produciendo a lo largo de la vida. El mismo Ramón y Cajal planteó la hipótesis de que los cambios observados en el comportamiento humano, tendrían probablemente un sustrato anatómico, hipótesis que en la actualidad se ha confirmado. Estos cambios son mayores en las primeras etapas de nuestra vida durante el desarrollo de circuitos neurales, pero en la madurez se siguen modificando nuestros contactos sinápticos a medida que aprendemos y establecemos memorias nuevas, que nos enfrentamos a situaciones fisiológicas diversas o cuando nuestro SN se ve sometido a enfermedades o lesiones. La capacidad de cambio continuo contribuye que el desarrollo y la organización del SN no esté marcado únicamente por la acción de los genes y que el cerebro de cada uno sea único.
Características estructurales y funcionales de la neurona
La variedad que presentan las neuronas en cuanto a forma y tamaño es tan amplia como las funciones que realizan. Sin embargo todas ellas presentan características comunes.
Como cualquier otra célula, la neurona está envuelta por la membrana neuronal, estructura que constituye el límite entre el interior celular y el medio que la rodea, permite a la célula funcionar como una unidad independiente. Las neuronas están rodeadas por el líquido intracelular compuesto por agua en la que están disueltas sales y muchas otras sustancias químicas, cuya concentración es diferente a ambos lados de la membrana para hacer posible el transporte de la información que es la función característica de las neuronas. La membrana está formada por una doble capa de fosfolípidos. La molécula fosfolipídica consta de dos zonas: la cabeza es la parte hidrófila (que tiene afinidad con el agua) mientras que las colas de los ácidos grasos son hidrófobas ( no tienen sitios de unión con el agua). La membrana plasmática tiene como característica esencial regular selectivamente el intercambio de sustancias entre el interior y el exterior celular. Para ello cuenta con diferentes moléculas proteicas insertadas en su doble capa lipídica que permiten que diversas sustancias atraviesen la membrana y de ellas van a depender muchas de las propiedades funcionales de la neurona. Algunas de las proteínas de la membrana forman canales que permiten el paso de sustancias, otras transmiten una señal al interior de la neurona cuando determinadas moléculas se unen a ellas en la superficie externa de la membrana, y otras las proteínas de la membrana actúan como transportadoras bombeando sustancias entre ambos lados de la membrana. La integración de la información que reciben a través de los contactos sinápticos y su transmisión a otras células, no podrían producirse si no fuera por la particular estructura y funcionamiento de la membrana neuronal.
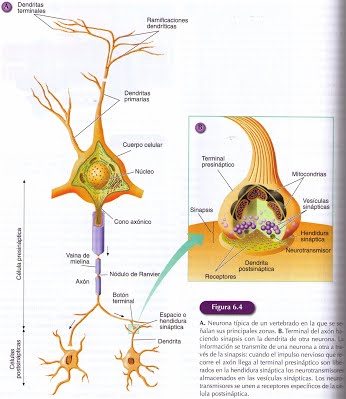
En la mayoría de las neuronas se puede distinguir tres zonas diferenciadas: el cuerpo celular, las dendritas y el axón.
Cuerpo celular o soma: Es el centro metabólico donde se fabrican las moléculas y realizan actividades fundamentales para mantener la vida y las funciones de la célula nerviosa. El interior de la célula está constituido por una sustancia gelatinosa, el citoplasma donde se localizan los mismos orgánulos que en otras células : el aparato de Golgim,los lisomas, una gran cantidad de mitocondrias, retículo endoplasmático rugoso y liso y diferentes estructuras fibrilares.
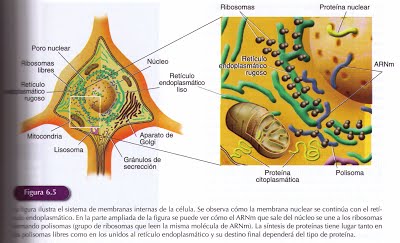
El cuerpo celular es la región que contiene el núcleo donde, al igual que en otras células, se localizan los cromosomasy el nucléolo que fabrica los ribosomas implicados en la síntesis de proteínas. Las proteínas son esenciales para las funciones de cualquier célula , pero las neuronas requieren además proteínas específicas para desarrollar su función especializada, la transmisión de información. Para sintetizar esas proteínas el cuerpo celular cuenta con gran cantidad de ribosomas y un complejo sistema de membranas formado por la continuación de la membrana nuclear con los tubos del retículo endoplasmático. Una zona de este sistema de membranas es el retículo endoplasmático rugoso, que se llama así, por el gran número de ribosomas que tiene adheridos. Las neuronas presentan tal acumulación de retículo endoplasmático rugoso que éste puede ser fácilmente visible en el microscopio. Se denomina sustancia de Nissl y mantiene una intensa actividad sintetizadora de proteínas.
Las proteínas sintetizadas tanto en los ribosomas unidos a las membranas del retículo endoplasmático rugoso como en los ribosomas libres penetran en los cuerpos de Golgi que son estructuras membranosas que empaquetan las proteínas para ser transportadas a diferentes destinos dentro de la neurona donde desempeñaran diversas funciones. Algunas pueden permanecer en el líquido celular y actuar como enzimas otras pueden incorporarse a la membrana celular donde actúan como canales o bombas para transportar sustancias a través de ella, o bien pueden ser destinadas a ser secretadas fuera de la célula como es el caso de los neurotransmisores.
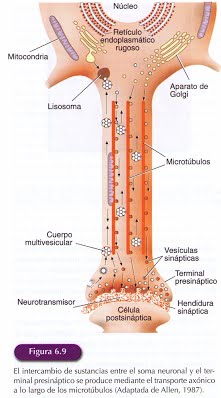
En el citoplasma de la neurona se localizan proteínas fibrilares o tubulares especializadas que constituyen el citoesqueleto. La organización de estas proteínas es fundamental para formar una matriz intracelular que determina la forma de la neurona, le da consistencia y proporciona un mecanismo de transporte moléculas en su interior. Si tenemos en cuenta que entre el soma y los terminales axónicos puede haber una distancia de un metro, como es el caso de las motoneuronas espinales que inervan nuestros pies, la eficacia de estos mecanismos de transporte de moléculas en el interior de la célula es fundamental.
 Los microtúbulos son los componentes más grandes del citoesqueleto y están directamente implicados en el transporte de sustancias en el interior celular. Los neurofilamentos o neurofibrillas son los elementos del citoesqueleto que más abundan en las neuronas. Su capacidad para retener el nitrato de plata permitió a Golgi desarrollar el método que hizo posible visualizar toda la neurona, lo que sirvió a Ramón y Cajal para establecer la Teoría Neuronal. Los neurofilamentos se encuentran entrelazados entre sí o con los microtúbulos, siendo está organización fácilmente susceptible de ser modificada bajo determinadas condiciones.
Los microtúbulos son los componentes más grandes del citoesqueleto y están directamente implicados en el transporte de sustancias en el interior celular. Los neurofilamentos o neurofibrillas son los elementos del citoesqueleto que más abundan en las neuronas. Su capacidad para retener el nitrato de plata permitió a Golgi desarrollar el método que hizo posible visualizar toda la neurona, lo que sirvió a Ramón y Cajal para establecer la Teoría Neuronal. Los neurofilamentos se encuentran entrelazados entre sí o con los microtúbulos, siendo está organización fácilmente susceptible de ser modificada bajo determinadas condiciones.
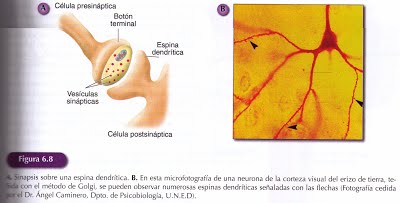
Las Dendritas: Son prolongaciones del soma neuronal con forma de árbol y constituyen las principales áreas receptoras de la información que llega a la neurona. La zona de transferencia de información de una neurona a otra es la sinapsis. La sinapsis tiene dos componentes el presinaptico y el postsináptico que señalan la dirección habitual del flujo de la información, que se produce desde la zona presinaptica hasta la zona postsináptica. Esta membrana dendrítica cuenta con un elevado número de receptores, que son las moléculas especializadas sobre las que actúan los neurotransmisores liberados desde otras neuronas.
La mayoría de las neuronas tienen varios troncos dendríticos que se ramifican varias veces mediante bifurcación, multiplicándose de esta manera el número de ramas dendríticas y en consecuencia el área que ocupa la neurona. La principal función de la esta ramificación dendrítica es incrementar la superficie de recepción de la información , ya que en toda la extensión del árbol dendrítico una neurona puede establecer miles de sinapsis al mismo tiempo. Las dendritas captan los mensajes y los conducen al cuerpo neuronal.
Algunas sinapsis se producen sobre pequeñas protuberancias de las dendritas denominadas espinas dendríticas. Las neuronas con escasas dendritas, cortas y poco ramificadas tendrán menos sinapsis que aquellas neuronas que su arborización sea extensa ya que permite recibir información desde un gran número de neuronas. Tanto la disposición y amplitud del árbol dendrítico, como el número de espinas, parecen ser susceptibles de ser modificados por una diversidad de factores ambientales constituyendo un ejemplo manifiesto de plasticidad neural.
 El axón: Es una prolongación del soma neural, generalmente más delgada y larga que las dendritas. Cada neurona tiene un solo axón y es la vía a través de la cual la información de propaga hacia otras células. Esta porción de la neurona también se denomina fibra nerviosa y su longitud varía entre algunas micras y varios metros, como es el caso de los axones de las neuronas motoras de las ballenas.
El axón: Es una prolongación del soma neural, generalmente más delgada y larga que las dendritas. Cada neurona tiene un solo axón y es la vía a través de la cual la información de propaga hacia otras células. Esta porción de la neurona también se denomina fibra nerviosa y su longitud varía entre algunas micras y varios metros, como es el caso de los axones de las neuronas motoras de las ballenas.
En el axón se pueden distinguir diferentes zona: un segmento inicial próximo al soma denominado cono axónico, el cual desarrolla una función integradora de la información que recibe la neurona, el axón propiamente dicho y el botón terminal, también denominado terminal del axón o terminal presinaptico. Del axón pueden surgir algunas ramificaciones colaterales, la ramificación primaria se produce ya en la zona distal, ramificándose después abundantemente en su terminación para transmitir la información al mayor número de neuronas.
En los extremos de las ramificaciones axónicas se encuentran los botones terminales, que se asemejan a un disco hinchado. Los botones terminales son el elemento presináptico de la sinapsis, pues a través de ellos el axón establece contacto con las dendritas o el soma de otra neurona o con otro tipo de célula para transmitir información. Contiene vesículas sinápticas con neurotransmisores que son liberados mediante exocitosis en el espacio extracelular cuando se producen unas condiciones muy determinadas.
El axón no cuenta con los orgánulos necesarios para que se pueda producir en él la síntesis de proteínas, por lo que estas moléculas han de ser constantemente suministradas desde el soma neuronal y transportados a través del axón. Los componentes del citoesqueleto del axón van a ser los encargados tanto del transporte de esas moléculas como de las diferentes sustancias del entorno celular que son captadas por el axón y transportadas hasta el soma.
Debido a la larga extensión que puede alcanzar el axón, la existencia de mecanismos de transporte eficaces es esencial para el correcto funcionamiento neuronal. Cuando el transporte se realiza desde el soma hasta el terminal se denomina anterógrado, cuando va desde el terminal sináptico hasta el cuerpo celular se denomina retrógrado. También distinguimos un transporte axónico rápido o un transporte axónico lento.
El flujo axónico rápido transporta los orgánulos celulares, como las mitocondrias, para atender a las necesidades energéticas y las vesículas que contienen neurotransmisores. El transporte rápido también se produce en dirección retrógrada(del terminal sináptico al cuerpo celular) para devolver al soma material procedente de los terminales sinápticos para su degradación o reutilización. Este material se empaqueta en el interior de estructuras multivesiculares que retornan al soma. Parece que un mismo microtúbulo puede hacerse cargo del transporte en ambos sentidos.
El transporte retrógrado no sólo sirve para eliminar desechos del terminal presináptico. Entre otras funciones está la de hacer llegar hasta el soma moléculas que son captadas por el terminal presináptico, como es el caso de los factores de crecimiento nervioso, sustancias que son liberadas desde la célula postsináptica , incorporadas por la célula presináptica y, mediante este transporte, conducidas hasta el soma neuronal para controlar la diferenciación neuronal durante el desarrollo del SN. La posibilidad de incorporar sustancias al terminal axónico para ser transportadas hasta el soma ha permitido el trazado de vías en el encéfalo. Las técnicas de trazado retrógrado permiten conocer la situación de los somas de las neuronas cuyos axones se encuentran sinaptando con las células de ese núcleo, para ello emplean sustancias que reaccionan en el interior de la neurona y cuyos productos de reacción pueden ser marcados para visualizar su trayectoria.
El flujo axónico lento interviene en el transporte de elementos del citoplasma. El citoplasma del axón es estructuralmente dinámico, ya que los elementos que lo componen están siendo constantemente reemplazados por materiales sintetizados y ensamblados en el soma que son transportados a lo largo del axón por el flujo axoplásmico lento. Esto es evidente durante el crecimiento y la regeneración de los axones, proceso que tiene lugar a una velocidad aproximada de 1mm al día, velocidad que coincide con los ritmos de transporte más bajos del flujo axónico lento.
Clasificación de las neuronas
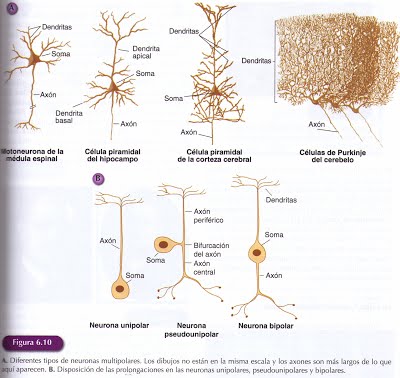 La clasificación más extendida hace referencia al número y disposición de sus prolongaciones.
La clasificación más extendida hace referencia al número y disposición de sus prolongaciones.
Neurona multipolar: Es el tipo neuronal más común y extendido en la escala zoológica. Además del axón, emergen del soma varias ramificaciones dendríticas. Según la longitud del axón se distinguen las multipolares de axón largo o Golgi tipo I,con cuerpo celular grande y varias dendritas muy ramificadas. Las células piramidales de la corteza cerebral y las células de Purkinge del cerebelo se encuentran en este grupo. Entre las multipolares de axón corto o Golgi tipo II, se encuentran muchas de las neuronas que realizan sus contactos con neuronas próximas incluidas dentro de la misma unidad funcional.
Neurona bipolar: Posee dos prolongaciones, un axón y una dendrita, que emergen de lugares opuestos del cuerpo celular. Estas neuronas se encuentran principalmente en los sistemas sensoriales, como es el caso de las células bipolares de la retina.
Neurona unipolar: Posee una sola prolongación que sale del soma. En las neuronas denominadas pseudounipolares, esta prolongación se divide en una porción que realiza la función de recepción de información propia de las dendritas y otra que realiza la función de conducción de información característica del axón. Estas neuronas son generalmente sensoriales. Las neuronas del sistema somatosensorial que detectan información táctil y nocioceptiva, las células amacrinas de la retina son este tipo de neuronas.
Las neuronas también pueden ser clasificadas según su función: las neuronas sensoriales captan la información del entorno, recibida a través de los órganos de los sentidos y la conducen al SNC, mientras que en las neuronas motoras o motoneuronas la dirección de la comunicación es opuesta, sus axones parten desde el SNC y llegan hasta los músculos con los que hacen sinpasis para que se produzca el movimiento.
Pero la mayoría de las neuronas que el SNC sólo establecen conexiones con otras neuronas, pueden ser de dos tipos: interneuronas o neuronas de circuito local y neuronas de proyección. Las interneuronas o neuronas de circuito local procesan información localmente, sus prolongaciones no salen de la estructura celular de la que forman parte. Las neuronas de proyección transmiten la información de un lugar a otro del sistema nervioso. Sus prolongaciones se agrupan formando vías que permiten la comunicación entre diferentes estructuras.
La Glía: Características y Tipos
En el SN nervioso existen además de las neuronas otro tipo de células que desempeñan un papel fundamental para mantener las neuronas en las condiciones óptimas que aseguren su funcionamiento y supervivencia.
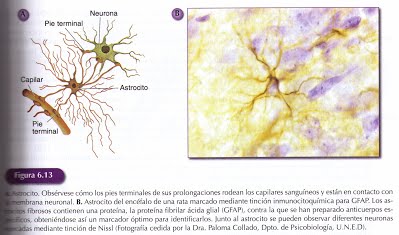 El complejo entramado neuronal se encuentra rodeado por las células gliales, neuroglia o glía. Son muy abundantes, más que las neuronas, y continúan dividiéndose en SN adulto. En el SNC existen varios tipos de células gliales: los astrocitos, los oligodendrocitos y la microglía. Fueron los estudios del histólogo Pío del Rio Hortega, los que permitieron distinguir los oligodendritos y la microglía. En el SNP las células de Shwann, aunque no se incluyen en la neuroglia, constituyen las células de soporte y realizan muchas de las funciones que desempeñan las células gliales en el SNC.
El complejo entramado neuronal se encuentra rodeado por las células gliales, neuroglia o glía. Son muy abundantes, más que las neuronas, y continúan dividiéndose en SN adulto. En el SNC existen varios tipos de células gliales: los astrocitos, los oligodendrocitos y la microglía. Fueron los estudios del histólogo Pío del Rio Hortega, los que permitieron distinguir los oligodendritos y la microglía. En el SNP las células de Shwann, aunque no se incluyen en la neuroglia, constituyen las células de soporte y realizan muchas de las funciones que desempeñan las células gliales en el SNC.
Los astrocitos
Son las células gliales más abundantes y se denominan así por su forma estrellada cuyas múltiples extensiones se dirigen en todas las direcciones desde el cuerpo celular. Algunos de los pies terminales de sus prolongaciones están en contacto con vasos sanguíneos, mientras otros envuelven la membranas somática y dendrítica de las neuronas.
Los astrocitos se presentan en dos formas principales: astrocitos fibrosos que se encuentran fundamentalmente en la sustancia blanca (constituida por haces de fibras nerviosas mielinizadas) y los astrocitos protoplasmáticos localizados en la sustancia gris, que es el tejido nervioso formado preferentemente por cuerpos celulares.
Los resultados de estudios recientes indican la existencia de una comunicación bidireccional entre astrocitos y neuronas, y por tanto la participación activa de éstos en el funcionamiento del SN. De los astrocitos se sabe:
Proporcionan soporte estructural. Se encuentran dispersos entre las neuronas formando una matriz que fija las neuronas y da consistencia al encéfalo
Regulan la transmisión sináptica. Mantienen alrededor de las neuronas las condiciones óptimas para que se produzca la transmisión de señales entre ellas. Aíslan a las sinapsis impidiendo la dispersión del neurotransmisor liberado desde los botones terminales en la hendidura sináptica, que intervienen en la recaptación y metabolismo de algunos neurotransmisores liberados por las neuronas en la sinapsis, que captan del espacio extracelular el potasio (K+) sobrante con lo que impide una acumulación que resultaría peligrosa para la función neural. Los astrocitos se pueden comunicar entre ellos y responden de manera compleja a la actividad neuronal. Además son capaces de liberar glutamato, neurotransmisor que modula la excitabilidad neuronal y la transmisión sináptica.
Intervienen en la reparación y regeneración del tejido nervioso Cuando las neuronas son destruidas, los astrocitos se activan y experimentan un aumento de tamaño. Para limpiar de desechos el cerebro, actúan ingiriendo y redirigiendo los restos de neuronas mediante fagocitosis y proliferan rápidamente para ocupar los espacios vacios formando un entramado de astrocitos. Desempeñan un papel restaurador liberando diversos factores de crecimiento que activan la regeneración de partes dañadas de las neuronas.
Recubren los vasos sanguíneos cerebrales y participan en el mantenimiento de la barrera hematoencefálica.
Suministra nutrientes a las neuronas. Los pies terminales de los estrocitos rodean los capilares cerebrales y las membranas neuronales para distribuir nutrientes, oxígeno, vitaminas y hormonas desde el sistema circulatorio hasta las neuronas y eliminar los productos de desecho. La glucosa que los astrocitos reciben desde los capilares es reducida a lactato que es incorporado por las neuronas para obtener energía.
Los oligodendrocitos y las Células de Schwann
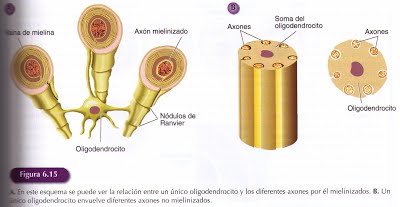 Los oligodendrocitos son pequeñas células gliales que emiten prolongaciones que se enrollan alrededor de los axones formando una densa capa de membranas que los envuelve denominada mielina.
Los oligodendrocitos son pequeñas células gliales que emiten prolongaciones que se enrollan alrededor de los axones formando una densa capa de membranas que los envuelve denominada mielina.
Esta vaina formada en su mayor parte por lípidos, constituye un aislante que mejora considerablemente la transmisión de impulsos nerviosos. La vaina de mielina no constituye una cubierta continua del axón, ya que se encuentra interrumpida cada milímetro por una zona de aproximadamente una micra, donde el axón queda al descubierto. Estas zonas se llaman nódulos de Ranvier.
Un único oligodendrocito puede mielinizar diferentes segmentos de un mismo axón o desarrollar prolongaciones que pueden formar segmentos de mielina en 20 o 60 axones diferentes. Los oligodendrocitos también pueden realizar una función protectora de los axones no mielinizados, envolviéndoles y manteniéndoles fijos por surcos formados por el soma de las células gliales.
Las células de Shwann realizan la mimas funciones en el SNP que las distintas células gliales en el SNC. Como los astrocitos se sitúan en el espacio interneural entre los somas neuronales y lo mismo que la microglía, fagocitan los desechos en caso de daño de los nervios periféricos. Una de sus principales funciones es formar la mielina alrededor de los axones del SNP. La mielina periférica se produce en fases tempranas del desarrollo al unirse las células de Shwann a los axones en crecimiento, las células de Shwann asociadas a ellos proliferan mediante división mitótica7 para mantener cubierto el axón donde se alinean a lo largo de toda su extensión. Cuando estas células de Schwann maduran, cada una de ellas desarrolla un único segmento de mielina para un único axón. Como ocurría en el SNC los axones amielínicos también son rodeados por células de Schwann que les separa unos de otros.
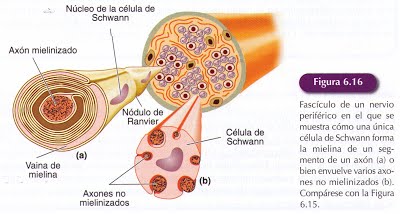 La formación de vainas de mielina se produce en varias fases, iniciándose cuando el axón queda completamente envuelto por el citoplasma de la célula de Schwann en una estructura que se denomina mesoaxón formada por la yuxtaposición de las láminas externas de la membrana de la célula de Schwann.
La formación de vainas de mielina se produce en varias fases, iniciándose cuando el axón queda completamente envuelto por el citoplasma de la célula de Schwann en una estructura que se denomina mesoaxón formada por la yuxtaposición de las láminas externas de la membrana de la célula de Schwann.
Posteriormente el mesoaxón se va alargando y va rodeando en espiral al axón. Para formar las finas capas concéntricas de la mielina madura, el citoplasma de la célula de Schwann se comprime. En el Sistema Nervioso Central la mielina se forma de la misma manera por los oligodendrocitos, aunque los axones se encuentran rodeados de un menor número de capas que los axones periféricos del mismo diámetro.
En nuestra especie el proceso de mielinización empieza en el segundo trimestre de vida fetal, aunque se intensifica después del nacimiento, continuando hasta la pubertad en algunas zonas del SN. La mielinización está estrechamente asociada al desarrollo de la capacidad funcional de las neuronas. Las neuronas cuyos axones han sido completamente mielinizados son capaces de realizar una rápida transmisión de impulsos, convirtiéndose en totalmente funcionales. Cualquier alteración que se produzca en la mielina tiene consecuencias importantes en el funcionamiento neuronal. Un ejemplo es la esclerosis múltiple que se caracteriza por la pérdida de mielina de los axones del SNC.
La Microglía
Son células pequeñas esparcidas por todo el SNC que se localizan entre las neuronas y los otros tipos de glía. En situaciones normales, el número de células de microglía es pequeño, no constituyendo más del 10% del total de células gliales. Pero cuando se produce una lesión o inflamación en el tejido nervioso estas células se vuelven reactivas, al igual que ocurre con los astrocitos. Como consecuencia de esta reactividad, las células microgliales cambian de apariencia lo que implica cambios morfológicos y funcionales. Proliferan rápidamente adoptan una forma ameboide y migrna a la zona del daño, donde fagocitan restos celulares, fragmentos de mielina o neuronas dañadas, participando en la reparación de la lesión.