Libros
Apuntes de Grado en Psicología - UNED
Primer curso
Psicología Social - Investigación en Psicología Social
Introducción al análisis de datos
Principios de aprendizaje y conducta
Segundo curso
Psicopatología. Volumen I - Psicopatología. Volumen II
Diseños de Investigación y Análisis de Datos
La Psicobiología es una disciplina de la Psicología. Su objetivo, por tanto, es el estudio del comportamiento humano y los procesos mentales que a él subyacen. Desde su origen, lo que diferenció a la Psicobiología del resto de disciplinas psicológicas y biológicas, fue el hecho de que dio máxima relevancia al sustrato biológico que hace posible el comportamiento y los procesos mentales, esto es, que encierra las claves de la naturaleza humana. Ese sustrato no es otro que el sistema nervioso. El sistema esculpido por la selección natural a lo largo de nuestra filogenia y que nos hace singulares. En la actualidad, ese atributo distintivo de la Psicobiología se desdibuja, pues la Psicología científica actual se encuadra en el mismo paradigma que, desde hace más de un siglo, preconiza la Psicobiología. Por ello, para poner de relieve su singularidad es conveniente contemplarla desde una perspectiva histórica.
No profundizaremos en más pormenores históricos que los estrictamente necesarios para lograr un punto de vista lo más claro posible del devenir de la Psicobiología. Por ello, tampoco entraremos aquí en el tema del dualismo mente-cuerpo, pero lo cierto es que la Filosofía mantuvo estancado bajo una férrea tutela el estudio de la naturaleza humana hasta finales del siglo XIX, momento extraordinario en el que, por un lado, se da a conocer al mundo la Teoría de la Evolución y, por otro, con la Teoría Neuronal, el estudio científico del sistema nervioso comienza a dar sus primeros frutos en la comprensión del comportamiento.
Tras El origen de las especies (1859), Darwin, publica, en 1871, El origen del hombre y la selección en relación al sexo, obra en la que pone de manifiesto que las diferencias en relación a las capacidades mentales del hombre y otros animales son sólo de grado y no de clase, es decir que, si bien el hombre tiene determinadas facetas sobresalientes en comparación con otros seres vivos, éstas se apuntan, ya existen en otros animales en mayor o menor medida según la proximidad filogenética a nosotros. Un año después, en 1872, en su libro La expresión de las emociones en el hombre y los animales, señala que el comportamiento es una característica biológica más que, al igual que cualquier otro rasgo biológico, está sujeto a la acción de la selección natural. Con estas tres obras, amén de las aportaciones que ellas mismas encierran, Darwin también da razón de ser al uso de modelos animales; una herramienta fundamental para el estudio del comportamiento y los procesos mentales.
Poco más tarde, en 1888, la Teoría Neuronal, planteada por nuestro más eminente científico, Santiago Ramón y Cajal, pone de manifiesto que el sistema nervioso está constituido por células, las neuranas, conectadas entre sí por contigüidad y no por continuidad. Su Teoría de la polarización dinámica de la neurona aporta también la primera explicación al funcionamiento y organización del sistema nervioso al proponer que las arborizaciones periféricas de la neurona serían las estructuras receptoras de las señales transmitidas por otras neuronas que, tras pasar por el cuerpo celular se enviarían por el cilindroeje hasta otras células nerviosas y órganos efectores. La información fluye, por tanto, desde los nervios periféricos a los centros nerviosos donde se procesa y parten las instrucciones para generar las respuestas, el comportamiento. El cerebro, de esta forma, deja de ser una maraña de fibras nerviosas sin orden alguno y se convierte en un órgano estructurado con vías que discurren de unas regiones a otras transportando información. Con ello, la Neurociencia comienza su fecunda andadura como nuevo pilar que sustenta el estudio científico del comportamiento humano y los procesos mentales que a él subyacen.
En este contexto, William James, en 1890, publica The Principies of Psychology, obra en la que, aunando las aportaciones de Darwin acerca de la función adaptativa del comportamiento y los avances en el estudio del sistema nervioso, trata de poner los cimientos de una Psicología científica que aborde su objeto de estudio a la luz del nuevo contexto que las ciencias naturales estaban creando. No obstante, aún se tardarían varias décadas en lograr el objetivo que W. James se propuso.
Un poco más tarde, en 1897, el fisiólogo ruso lván Paulov, formula la Ley del reflejo condicional, base de lo que hoy conocemos como condicionamiento clásico. En 1903, Edward Thorndike, discípulo de W. James, descubre la Ley del efecto que propició la formulación, más tarde, del condicionamiento operante de Skinner. Con estos mimbres, en 1913, John B. Watson, funda el Conductismo y, con él, la Psicología científica. Bajo el paradigma conductista, el estudio científico del comportamiento se centra en dos variables principales: el estímulo y la respuesta (E-R). La respuesta es la conducta que, de esta forma, pasa a ser un hecho positivo, objetivo, perceptible por los sentidos, verificable y susceptible de ser cuantificado, quedando excluidas aquellas manifestaciones cuyo conocimiento sólo es alcanzable por medio de la introspección y no pueden, por tanto, ser verificadas por otro observador.
No obstante, la Psicología científica comienza su andadura más como un conjunto de corrientes simultaneas dirigidas a dar una explicación del comportamiento humano que como una disciplina cohesionada. Conductistas, neoconductistas, cognitivistas y psicobiólogos investigan el estudio del comportamiento humano y los procesos mentales desde el ámbito científico pero bajo distintos paradigmas.
Para los conductistas, el estudio del comportamiento debía reducirse al paradigma E-R; su objeto de estudio eran las respuestas, el comportamiento mostrado ante la exposición a un estímulo; el organismo (el cerebro) era como un papel en blanco 4 en el que el esfuerzo, la cultura y la educación representaban los auténticos y únicos determinantes del comportamiento humano y, por tanto, del futuro de las personas.
«Dadme una docena de niños sanos, bien formados, y mi mundo especificado donde criarles, y garantizo que tomaré a cualquiera de ellos al azar y le educaré para que llegue a ser cualquier tipo de especialista que yo decida: médico, abogado, artista, comerciante y, sí, incluso pordiosero y ladrón, cualesquiera que sean sus dotes, inclinaciones, tendencias, habilidades, vocaciones y la raza de sus antepasados», -- afirmaba Watson, en su obra Behaviorism (1930).
Por su parte, para los neoconductistas, que se centraban principalmente en el aprendizaje, el organismo debía pasar a ser un elemento activo de la ecuación que diese explicación al comportamiento (E-0-R), pero esa implicación no pasaba de considerar al organismo más que un mero receptáculo de determinadas variables no observables (los procesos mentales) intercaladas entre el estímulo y la respuesta. Más tarde, en los años sesenta del pasado siglo, los cognitivistas enfocan el estudio de los procesos mentales bajo un modelo cibernético considerando irrelevante el soporte físico donde esos procesos ocurrían (el SN). Hoy, afortunadamente ese panorama ha cambiado, la Neurociencia Cognitiva es un buen ejemplo de ello y se puede decir que ya todas las disciplinas de la Psicología científica abordan sus objetivos propios dentro del marco que la Psicobiología comenzó a crear a principios del pasado siglo.
El término Psicobiología es acuñado, en 1914, en el libro An Outline of Psychobiology (Un esquema de Psicobiología), escrito por el psicólogo estadounidense y presidente de la American Psychological Association, Knight Dunlap. Su objetivo fue crear una pequeña introducción para sus alumnos con el fin de «ayudar a los estudiantes de Psicología que no han realizado cursos de Biología, a adquirir las bases morfológicas y fisiológicas directamente relacionadas con la Psicología». Tres años después, en 1917, este «Outline », lo desarrollará Dunlap fundando la revista Psychobiology, con una finalidad que deja claro lo que la Psicobiología pretende: «publicar las investigaciones en ciencia básica que tengan en cuenta la interconexión de las funciones mentales y fisiológicas». En 1921, la revista se fusionará con Journal of Animal Behavior, fundada en 1911, por el primatólogo estadounidense Robert Yerkes, en la que se publicaban los resuItados de investigaciones sobre sensación, percepción, aprendizaje y Etología. Nace así, Journal of Comparative Psychology que, en 1942, pasa a llamarse Journal of Comparative and Physiological Psychology.
Finalmente, en 1983, como consecuencia de la irrupción del campo multidisciplinar de la Neurociencia, la revista se dividió en Journal of Comparative Psychology y Behavioral Neuroscience que, en palabras de la actual editora de la publicación, Rebecca D. Burwell, es la descendiente directa de Psychobiology fundada por Dunlap. Unos años antes, en 1968, comienza a editarse Developmental Psychobiology, destina a recoger las investigaciones que sobre el desarrollo del comportamiento tienen los factores epigenéticos en la etapa perinatal y la primera infancia.
La Psicobiología sigue, por tanto, el paradigma E-0-R, pero carga de contenido cada uno de sus elementos. Entre el estímulo y la respuesta no se intercala ni una «pizarra en blanco» ni un mero receptáculo de variables no observables como definían los neoconductitas al organismo, sino el sistema nervioso fruto de la historia filogenética de nuestra especie, en el cual se lleva a cabo el procesamiento de la información estimular y los procesos mentales que hacen posible el comportamiento humano. La Psicobiología no sólo incorpora a su objeto de estudio las aportaciones de la Neurociencia y la Psicología científica, también recoge las realizadas desde la genética de la conducta y otras disciplinas biológicas.
Sin embargo, toda la actividad generada a finales del siglo XIX y comienzos del XX, se ve en cierta medida enlentecida (fundamentalmente en Europa) por las dos Guerras Mundiales y, en nuestro caso, también por la Guerra Civil española. El exilio de investigadores y las subsiguientes etapas de postguerra, ralentizaron el desarrollo de la Psicología científica en Europa y en especial en España. En nuestro país, tras la Guerra Civil, a pesar del Premio Nobel en Fisiología y Medicina concedido a Cajal en 1906, de la intensa actividad científica y académica que ello supuso con la creación de instituciones como la Junta de Ampliación de Estudios e Investigaciones Científicas, de la creación del Laboratorio de Investigaciones Biológicas, luego Instituto Cajal; de la fundación de revistas como Trabajos del Laboratorio de Investigaciones Biológicas; de que, ya en 1902, el neurólogo Luis Simarro ocupase, en la Universidad Central de Madrid, la primera Cátedra de Psicología Experimental de España y de que, en 1928, se crearan los primeros institutos psicotécnicos en Madrid y Barcelona, a pesar de todo ello, tras 1939, los estudios de Psicología vuelven de nuevo al ámbito de la Psicología escolástica dentro de la Licenciatura de Filosofía y Letras. En él permanecerán oficialmente hasta los años ochenta del pasado siglo en los que, finalmente, se crean las primeras Facultades de Psicología y departamentos de Psicobiología. Consecuencia, en gran parte, de que, desde los años cincuenta del pasado siglo, paulatinamente la enseñanza universitaria de la Psicología comenzara a retornar al ámbito de la Psicología científica, gracias al impulso aportado por jóvenes investigadores formados fuera de España, como Mariano Yela, José Luis Pinillos, Manuel Úbeda, Miguel Siguán, Francisco Secadas o José Forteza, entre otros.
La Explicación de la Conducta
Para la Psicobiología, la conducta y los procesos mentales son una propiedad biológica que, como el resto de características de los seres vivos, han sido modelados por la selección natural, es decir, son reflejo de la evolución y junto con los otros dos elementos del paradigma E-O-R, el estímulo y el organismo, forma lo que se denomina un complejo adaptativo.
Las características de este complejo adaptativo varían entre las especies y en menor medida de unos individuos a otros, ya que dependen de dos factores. El primero de ellos es el filogenético y hace referencia a la historia evolutiva que ha experimentado la especie. El segundo es el factor ontogénico y recoge las circunstancias en las que se ha desarrollado la vida del individuo desde el momento de su concepción.
El factor filogenético está representado por el acervo genético de la especie a la que pertenece el animal, en el cual se recogen los logros adaptativos de sus predecesores que han resultado ventajosos para la supervivencia de la especie a lo largo de su devenir y que se plasman en las características de las estructuras y órganos receptores, de los sistemas que integran la señal estimular y de los sistemas efectores encargados de emitir las respuestas. A las adaptaciones conseguidas a lo largo de la filogenia que recoge el acervo genético de la especie y que capacitan a cualquier miembro de la misma para recibir un determinado espectro estimular, procesar de determinada forma esta información y emitir una respuesta conductual, se les denomina causas lejanas del comportamiento. Son, por tanto, las responsables de las diferencias que existen entre las especies, causantes, por ejemplo, de que las abejas reaccionen a la luz ultravioleta, de que nuestra especie pueda comunicarse a través del lenguaje o de que las aves vuelen ante la presencia de un depredador.
El segundo factor involucrado en las características del complejo adaptativo es el factor ontogénico. Con él se quiere señalar que, si bien la filogenia marca un patrón general que identifica a cada ser viviente como perteneciente a una especie o a otra, dentro de las distintas especies cada uno de sus miembros es diferente a cualquier otro. Ello es debido, en gran medida, a que la dotación genética de cada individuo es el resultado de una combinación única del genoma de la población que lo hace singular, tanto en sus rasgos físicos como psicológicos. Sin embargo, cuando se indica que un carácter morfológico, fisiológico o conductual, depende o está controlado genéticamente, no hay que interpretarlo siempre en un sentido determinista. Las características únicas de cada individuo son consecuencia también de la interacción que se establece entre su genotipo y el ambiente; la importancia que ejerza cualquiera de estos dos factores dependerá del rasgo estudiado. A los factores ambientales que actúan modulando la expresión génica se les denomina factores epigenéticos. Sus efectos sobre el sistema nervioso pueden tener un mayor o menor grado de reversibilidad. Los efectos menos reversibles están asociados a determinados periodos de máxima susceptibilidad del sistema nervioso, los denominados periodos críticos que, generalmente, se circunscriben a la etapa perinatal. Un ejemplo de ello es la acción que ejercen las hormonas sexuales en las primeras etapas del desarrollo postnatal de diversas aves y mamíferos, alterando morfológica y fisiológicamente algunas regiones del sistema nervioso involucradas en la conducta sexual y otro tipo de comportamientos. Los efectos más reversibles están relacionados con una propiedad muy importante del sistema nervioso: la plasticidad neuronal, que es la capacidad que tienen las neuronas de experimentar cambios en su morfología y fisiología frente a distintas situaciones ambientales. Esta propiedad ha tenido gran importancia a lo largo de la filogenia al favorecer la aparición de procesos tan importantes como el aprendizaje y la memoria que, a su vez, han permitido el desarrollo de sistemas nerviosos que respondan de forma más flexible y eficiente a los retos ambientales.
Finalmente, a los factores indicados hay que añadir aquellos otros que intervienen en la causación inmediata del comportamiento. Entre éstos se encuentran los mecanismos por los que las diferentes energías estimulares son captadas por los receptores sensoriales y se integran dentro del sistema nervioso central (SNC), (procesos de sensación y percepción ); el cómo dicha representación del entorno produce cambios en el estado interno del organismo (procesos de motivación, emoción y aprendizaje), y de qué manera esos cambios fisiológicos en el medio interno influyen en la forma en que el organismo interactúa con su ambiente al desplegar el comportamiento.
El conjunto de factores responsables de la singularidad conductual de cada individuo constituye las causas próximas del comportamiento. Son, por tanto, las responsables de que las características generales de la especie se expresen de modo particular en cada individuo, aportando la diversidad imprescindible para la supervivencia y evolución de las especies.
Por todo ello, la Psicobiología considera a la conducta como el conjunto de manifestaciones públicamente observables reguladas por el sistema nervioso, mediante las cuales el organismo, como un todo, en respuesta a un estímulo interno o externo, se relaciona activamente con el medio ambiente de la forma que determine su devenir filogenético y ontogénico.
La Psicobiología, además, trata de dar una explicación en términos fisicalistas, no sólo de la conducta, sino también de los procesos mentales que la hacen posible. La mente está ineludiblemente ligada al organismo, es un producto de su actividad neural , consecuencia de la acción de la selección natural y, por tanto, dependiente enteramente del sustrato biológico que la genera. Como indica el neurofisiólogo Rodolfo Llinás, en su libro, El cerebro y el mito del yo (2003):
«El primer paso fundamental para explorar, desde un punto de vista científico la naturaleza de la mente es rechazar la premisa de que, ésta apareció súbitamente como resultado de una "intervención espectacular''. La naturaleza de la mente debe entenderse con base en su origen, en el proceso de su desarrollo, que emana del perenne mecanismo biológico de ensayo y error. La mente, o lo que llamaré "el estado mental'; es el producto de los procesos evolutivos que han tenido lugar en el cerebro de los organismos dotados de movimiento. Esta evolución cerebral se presentó de manera paulatina, desde las formas más primitivas hasta las más altamente evolucionadas por tanto, el examen de las bases científicas de la mente requiere una perspectiva evolutiva rigurosa ya que es a través de este proceso como se generó la mente».
Por ello, cualquier intento de explicación de la mente como proceso independiente del organismo está abocado al fracaso.
Los fenómenos mentales son fenómenos cerebrales y uno de los objetivos de la Psicobiología es identificar también los sistemas neurales cuya actividad específica es mental (afectiva, perceptiva, intelectual o volitiva) y explicar dicha actividad mental.
El comportamiento humano tiene múltiples condicionantes algunos de los cuales fueron abordados en un primer momento por disciplinas ajenas a la Psicología, tales como la Genética, la Etología o la Neurociencia. La Psicobiología ha ido integrando todas esas aportaciones creando un nuevo marco de análisis del comportamiento e incorporando a su actividad investigadora todas aquellas técnicas y objetivos particulares de esas disciplinas que le permitan explicar plenamente su objeto de estudio. El resultado de esa síntesis ha sido la aparición paulatina de nuevas disciplinas que están aportando un importante cuerpo de conocimientos que permiten hoy conocer y explicar mejor el comportamiento humano, a la vez que abren nuevos horizontes para su estudio.
Las causas lejanas de la conducta son el ámbito de estudio de la Psicología evolucionista. Esta disciplina parte de la base de que para comprender y explicar plenamente la conducta humana y los procesos mentales que la hacen posible, debe tenerse en cuenta su historia evolutiva. Para ello, recurre al análisis comparado de la conducta y los procesos mentales en el contexto de las relaciones filogenéticas de las especies. Este tipo de estudios involucra a otras disciplinas cuyos objetivos trascienden a los de la Psicobiología por no circunscribirse sólo a nuestra especie sino al conjunto de animales, pero que, por ello mismo, también los abarca. Es el caso de la Etología, la Ecología del Comportamiento y la Sociobiología, que en el contexto de la Teoría Sintética de la Evolución, consideran que la conducta, como cualquier otra característica de los seres vivos, es fruto de la selección natural y tratan de poner de manifiesto esta circunstancia. Sus estudios se centran en la observación y evaluación de la conducta de diversas especies animales en condiciones naturales o lo más parecido posible a ellas, tratando de establecer relaciones entre medidas biológicas y conductuales, como es el caso de la Etología, analizando el control neuronal de dicho comportamiento, que es de lo que se encarga la Neuroetología, o estudiando determinadas estrategias conductuales en relación con sus implicaciones ecológicas y evo lutivas, competenc ia ésta de la Ecología del Comportamiento. La Sociobiología, por su parte, estudia las bases biológicas del comportamiento social y es resultado de la confluencia de la Genética de Poblaciones, la Ecología y la Etología. En 1975, Edward Wilson, fundador de esta disciplina, la define como: «el estudio sistemático de la base biológica de todas las formas de comportamiento social en toda clase de organismos». Su propósito es formular leyes generales de la evolución y biología del comportamiento social que puedan explicar esta faceta del comportamiento humano.
En un plano más próximo, en cuanto a la causalidad del comportamiento, se encuentra la Genética de la Conducta. El objetivo de esta disciplina podemos decir que se circunscribe al ámbito psicobiológico aunque, cuando nace, ni la Psicobiología ni la Psicología científica existían. En 1869, Francis Galton, a raíz de la publicación de la obra de Darwin, El Origen de las especies, da a conocer los primeros estudios sobre la herencia de la capacidad mental humana, en el libro El genio hereditario: examen de sus leyes y consecuencias, en el que pone de manifiesto la existencia de una correlación entre la proximidad familiar y la capacidad mental, sentando de esta forma las bases de lo que luego sería la Genética de la Conducta. Como indica el genetista de la conducta Robert Plomin, profesor de genética del comportamiento del Instituto de psiquiatría de Londres, en esta disciplina se pueden distinguir tres tipos de enfoques: el centrado en el gen, que comienza con un efecto genético sencillo y estudia su influencia sobre la conducta. El enfoque centrado en la fisiología , que se concentra en los intermediarios fisiológicos entre genes y conducta, pudiendo tomar dos direcciones, una dirigida a averiguar los genes responsables de dichos intermediarios fisiológicos y otra a estudiar el efecto que éstos tienen sobre la conducta. El tercer enfoque comienza en la conducta y trata de averiguar en qué medida las influencias genéticas o ambientales afectan a la misma.
Los condicionantes genéticos son modulados por los factores epigenéticos. Del estudio de estas interacciones y de las consecuencias que ellas tienen sobre la conducta se encarga la Psicobiología del desarrollo. Gracias a los trabajos realizados dentro de esta disciplina se han descubierto, por ejemplo, las alteraciones que se producen en el desarrollo de la corteza cerebral cuando existe una deficiencia de hormonas tiroideas en la etapa prenatal o los efectos de la privación sensorial y el aislamiento social.
Los condicionantes inmediatos de la conducta son competencia de disciplinas como la Psicología Fisiológica que, mediante la intervención sobre el SN, estudia las bases biológicas del comportamiento, intentando explicar los cambios en el organismo durante el desarrollo de una conducta; la Psicofarmacología, que se centra en el estudio de las características estimulares de los fármacos y las drogas de abuso, así como en la influencia que sobre ese efecto tienen las variables ambientales; la Psicofisiología que estudia, sin manipular el SN, los cambios fisiológicos producidos en humanos ante determinadas situaciones o tras la presentación de distintos estímulos o la Psiconeuroendocrinología, disciplina que centra su interés en conocer los mecanismos por los que las hormonas afectan al SN, la conducta y a los procesos psicológicos, y cómo éstos a su vez pueden influir sobre el sistema endocrino.
En el ámbito clínico, como una confluencia entre la Psicología y la Neurología, y con el fin, en origen, de estudiar los efectos que las lesiones del SNC provocaban sobre la conducta y los procesos mentales, surgió la Neuropsicología. Esta disciplina se centra, dentro del contexto clínico, en conocer qué estructuras del sistema nervioso participan en los procesos psicológicos humanos como el aprendizaje, la memoria, el lenguaje, los procesos cognitivos, el procesamiento de la información, la resolución de problemas o los procesos emocionales. Recientemente, centrada en los procesos cognitivos, surge la Neurociencia Cognitiva.
La ciencia constituye un cuerpo organizado de conocimientos que proporciona un control práctico sobre el mundo físico e influye de manera importante en la concepción que el hombre tiene del mundo que le rodea. La Psicobiología es una ciencia empírica y comparte, por tanto, la visión científica del mundo, el método científico y los objetivos generales de la ciencia. Su meta es explicar la conducta y los procesos psicológicos en términos fisicalistas, como el resultado de la actividad del sistema nervioso.
El conocimiento científico es aquel que se obtiene mediante la utilización del método científico, una herramienta sencilla, pero potente. Gracias a él, hoy en día accedemos a casi cualquier faceta de la cultura con tan solo un clic; se mejoran cosechas y condiciones de vida; se erradican, curan y palían enfermedades, y se avanza para entender la etiología de muchas otras. Si la humanidad en el último siglo ha pasado de mil seiscientos millones de habitantes a más de siete mil millones, se debe, en buena parte, a la actividad científica y tecnológica que deriva de ella. Lo que depare el futuro a la humanidad depende también, en gran medida, del conocimiento preciso y objetivos que tengamos de nuestra propia naturaleza, de sus posibilidades y limitaciones. Ese es también el objetivo de la Psicobiología.
Hablar de método en ciencia es hablar del método hipotético-deductivo, método que hace referencia al planteamiento y verificación de hipótesis y, a partir de ahí, a la formulación de leyes y establecimiento de teorías. El método científico parte de la observación, entendida ésta como la información suministrada por la experiencia. Es la fuente principal de la que se nutre la ciencia, es decir, los hechos que trata de explicar y, como veremos más adelante, el control y aceptación de dichas explicaciones. Pero la observación no es el acopio de datos. Como decía Darwin: «la razón de ser del observar no reside en meramente recoger y acumular observaciones sino en buscar y sacar a la luz cierto orden existente en los hechos». Como primer paso para explicar los problemas planteados por la observación, el científico recurre a formular hipótesis que son conjeturas que propone para dar cuenta de los hechos observados. La hipótesis debe ser verosímil, guardar alguna relación con conocimientos previos alcanzados por la ciencia y ser susceptible de contrastación empírica.
La contrastación empírica de las hipótesis se realiza a través de los enunciados deducidos a partir de las hipótesis (de ahí que el método científico sea llamado también método hipotético-deductivo). La hipótesis no es sometida a contrastación empírica, lo que se contrasta son casos concretos deducidos a partir de la hipótesis. A medida que aumenta el número de casos favorables de la contrastación, mayor será el apoyo empírico con que cuenta la hipótesis y, en consecuencia, aumenta su grado de verosimilitud o de probabilidad. Este hecho pone en evidencia una de las características de las ciencias empíricas y es que éstas no proporcionan un conocimiento en términos absolutos, como ocurre con las ciencias formales (ej. las matemáticas), sino en términos relativos, de probabilidad.
En las ciencias empíricas la comprobación de una hipótesis involucra la experiencia, pero no son necesariamente experimentales. Para probar empíricamente las hipótesis, el investigador puede optar por la observación o por la experimentación. Esta última supone la modificación deliberada de las condiciones de contrastación derivadas de la hipótesis para la comprobación de la misma, mientras que la contrastación observacional se limita a registrar variaciones.
El método científico permite llegar a conclusiones a partir de los datos obtenidos por la observación y la experimentación. Esta conclusión es lo que constituye la ley científica, que supone relacionar los datos obtenidos en la contrastación con la hipótesis planteada, permitiendo explicar las relaciones que se dan entre los hechos observados. De esta forma, las hipótesis representan la primera aproximación al conocimiento científico, siendo las leyes derivadas de ellas la explicación científica a un hecho de observación. Cuando una serie de leyes pueden agruparse para explicar fenómenos completamente diversos, permitiendo una comprensión unificada de los mismos, de carácter más general que el descrito por cada una de esas leyes, aparece una teoría.
Por tanto, la explicación científica se articula de una manera extremadamente flexible, constituyendo un cuerpo de conocimientos obtenidos mediante el método hipotético-deductivo. En ciencia no hay dogmas porque el conocimiento científico siempre es provisional y susceptible de revisión cuando nuevos datos u observaciones reemplazan a los planteamientos existentes. Derivada de esa característica y de la imposibilidad de universalizar las leyes y teorías científicas, el epistemólogo Karl Popper, propuso el falsacionismo como forma de contrastación de las hipótesis y las teorías. Bajo esta doctrina, la forma más eficiente de corroborar hipótesis y teorías es intentando refutarlas mediante contraejemplos. Si no es posible refutarlas, las hipótesis y las teorías quedarán corroboradas y podrán ser aceptadas provisionalmente por las evidencias empíricas, pero nunca serán verificadas 9 .
Bajo este contexto, la Psicobiología trata de explica r la conducta humana a través del funcionamiento del sistema nervioso y del organismo en general. De una forma esquemática podemos señalar que las estrategias para explicar la conducta y los procesos mentales que a él subyacen, implican co ntrastaciones experimentales y observacionales. Dentro de las primeras existen dos grandes estrategias:
- la intervención conductual, en la que la conducta actúa como variable independiente (VI) y el organismo (el SN) como variable dependiente (VD) y
- la intervención somática, en la que se toma a la conducta como variable dependiente siendo el organismo (el SN) la variable independiente.
En la intervención conductual se manipula el ambiente para tratar de producir alguna modificación conductual concreta (variable independiente), evaluándose el efecto que dicha manipulación ha tenido sobre el organismo (variable dependiente). Lógicamente la forma de propiciar los cambios conductuales que se estimen oportunos es actuando sobre el ambiente en el que se desenvuelve el sujeto experime ntal, es decir, controlado las distintas variables estimulares a las que el sujeto es expuesto a través de sus sentidos: contacto, temperatura, presión, iluminación, olores, sonidos, sabores. Estos estímulos se pueden proporcionar directamente a través de, por ejemplo, imágenes u olores específicos, o bien a través de, por ejemplo, manipulación de objetos, la presencia de individuos del mismo o distinto sexo, cambios el ciclo de luz-oscuridad, la disponibilidad de comida y bebida, etc. De esta forma, controlando el comportamiento del anima l (variable independiente) evaluaremos su efecto sobre las variables (dependientes) que consideremos oportunas del SN: estructuras invo lu cradas en determinados comportamientos, procesos fisiológicos afectados, etc.
Un ejemplo de intervención conductual nos lo proporcionan toda la serie de investigaciones encaminadas a contrastar la hipótesis formulada por Donald Hebb, en 1949, conocida como la teoría de la asamblea celular, en la que planteaba que la actividad neuronal repetida como consecuencia de una experiencia reiterada provocaría cambios en las neuronas involucradas que potenciarían su conexión formando redes o asambleas neuronales que favorecerían respuestas más eficientes al exponerse de nuevo a esa experiencia. Tuvieron que pasar casi veinte años hasta que las técnicas permitieron poner a prueba esa hipótesis. Ésta comenzó en los años sesenta del pasado siglo, de la mano el grupo liderado por el psicólogo estadounidense Mark R. Rosenzweig, en Berkeley. Estos investigadores realizaron una serie de trabajos en los que pusieron de manifiesto que el entrenamiento formal y la experiencia informal adquirida por la exposición a ambientes variados (variables independientes), provocaban cambios medibles en la neuroquímica y neuroanatomía del cerebro de roedores (variables dependientes). Comprobaron, por ejemplo, que aquellos animales que habían sido entrenados en la resolución de distintos tipos de test comporta mentales (VI) presentaban una mayor actividad de la enzima acetilcolinesterasa (AChE) en la corteza cerebral (VD), que los que no habían sido entrenados en este tipo de test, además existía una correlación positiva entre la cantidad de AChE y la dificultad del test. Asimismo, descubrieron que aquellos animales estabulados en ambientes que generaban distintas oportunidades de aprendizaje informal (ambiente enriquecido) (VI) presentaban también más cantidad de AChE y mayor peso de la corteza cerebral (VD ) que los criados en las condiciones habituales de estabulación (ambiente empobrecido) (VI). Trabajos posteriores han extendido el número de variables que se ven modificadas en el SN de mu chas especies de aves y mamíferos como consecuencia de la experiencia y la actividad del SN, corroborando, de esta forma, la hipótesis de Hebb.
En la intervención somática se producen alteraciones sobre el SN, a través de, por ejemplo, lesión de estructuras cerebrales, inyección de fármacos, etc, que pasarían a ser, por tanto, variables independientes, pues son las que controlamos para evaluar los ca mbios en la conducta del sujeto que es la variable dependiente.
Un ejemplo de este tipo de intervención lo tenemos en los trabajos que han ido descubriendo el dimorfismo sexual de distintas regiones del SNC, causado por el efecto que las hormonas sexuales ejercen sobre ellas en el periodo perinatal. En 1959, Phoenix, Robert W. Goy, Arnold A. Gerall, y William C. Young, en la Universidad de Kansas, demuestran que la administración prenatal de testosterona (VI) masculinizaba el comportamiento de las cobayas hembra adultas (VD) por su efecto sobre las regiones del SNC involucradas en el conducta sexual. Posteriormente, estos resultados se replicaron en monos rhesus, mostrando, además, que el comportamiento de juego de los monos hembra (VD) tratados prenatalmente con la testosterona (VI) también se masculinizaba.
Desde entonces, numerosos trabajos han ido poniendo de manifiesto diferencias de sexo en distintos núcleos del SNC, es el caso, por ejemplo, de los realizados por el grupo de Roger Gorski, en la Universidad de California en Los Ángeles, que en 1978, descubren la existencia de un núcleo del área preóptica (un área involucrada directamente en la exhibición del comportamiento sexual del macho en muchas especies) que es mucho mayor en machos que en hembras; le denominaron por ello, núcleo sexodimófico del área preóptica (NSD). Comprobaron también, que esas diferencias dependían de la presencia de esteroides sexuales durante el período perinatal. En paralelo, pusieron de manifiesto que los roedores hembra a los que se les administraba testosterona al comienzo del desarrollo (VI), mostraban el comportamiento de monta típico de los machos (VD), mientras que los machos privados de la testosterona mostraban el comportamiento de lordosis típico de las hembras (VD).
Junto a estas estrategias, existe una tercera forma de abordar el estudio de la conducta que implica una contrastación observacional. Ésta se utiliza cuando no existe posibilidad de usar una contrastación experimental. En Psicobiología, dado que en ocasiones, al investigar con sujetos humanos o con otros animales, no es posible el control de las variables independientes, se recurre a la contrastación observacional para llevar a cabo la verificación o falsación de una hipótesis, siendo la aproximación correlacional, es decir, la observación de covariaciones entre medidas biológicas y conductuales, la forma de dar explicación de la hipótesis de partida. Un buen ejemplo de aproximación correlacional lo tenemos en el famoso caso Phineas P. Gage (1823-1860), uno de los primeros casos documentados de daño cerebral y, por tanto, histórico en el estudio de las bases biológicas del comportamiento. Gage fue un obrero de ferrocarriles que, en 1848, sufrió un accidente que le causó importantes daños en el cerebro, específicamente en parte del lóbulo frontal. Éstos se produjeron como consecuencia de una explosión que provocó que una barra de metal saliese disparada atravesando el cráneo de Gage para aterrizar, finalmente, a casi 30 metros de distancia. La barra de un metro de largo y más de 3 cm de diámetro entró en su cráneo por la mejilla izquierda y salió por la parte superior atravesando la corteza cerebral anterior. Sorprendentemente, Gage se mantuvo consciente en todo momento y la crónica de los médicos del pueblo relata que a los pocos minutos del accidente fue capaz de hablar y de caminar. Unos meses después parecía totalmente recuperado y volvió a su trabajo, sin embargo, aquéllos con los que se relacionaba notaron un cambio importante en su personalidad y su conducta. El movimiento y el habla no se vieron afectados, pero pasó de ser una persona sensible y respetuosa a ser maleducada y grosera que, además, mostraba conductas desinhibidas, impulsivas e imprudentes. El accidente, por tanto, provocó varias lesiones incontroladas en el cerebro de Gage y toda una serie de alteraciones conductuales. Obviamente, en este caso no existe un control de las variables como existiría si el trabajo se realizase con animales de experimentación, en los que podemos provocar una lesión de forma muy precisa en magnitud y localización, como hemos descrito anteriormente a la hora de explicar la intervención somática. No existen, por tanto, variables dependientes e independientes, sino un daño cerebral en una región amplia de la corteza cerebral y la alteración conductual subsiguiente que permitió a los investigadores, a través de una aproximación correlacional, involucrar a los lóbulos frontales en aspectos relacionados con la personalidad, la emoción y la interacción social.
En general, el estudio de personas con daño cerebral ha permitido a los neuropsicólogos descubrir la función de muchas regiones del sistema nervioso central y los mecanismos involucrados en los procesos psicológicos superiores. Las áreas de Broca y Wernicke, cuyo daño es responsable del síndrome afásico, son otros dos ejemplos en los que la contrastación observacional, a través de la aproximación correlaciona! permitió a Paul Broca, en 1861 y Carl Wernicke, en 1874, descubrir, poco después del caso de Phineas P. Gage, que el daño de una zona del lóbulo frontal izquierdo (área de Broca) y una región del lóbulo temporal izquierdo (área de Wernicke) eran los responsables del síndrome afásico y, con ello, la participación de esta dos regiones en la producción y comprensión del lenguaje.
La Neuropsicología y la Neurociencia Cognitiva se han ido paulatinamente liberando de su dependencia de los traumatismos encefálicos y accidentes cerebrovasculares para el estudio de los procesos psicológicos superiores y el sustrato neural que los sustenta. Primero, con el desarrollo de las técnicas de electroencefalográfia (EEG) y potenciales evocados (PE) y, desde los años 80 del pasado siglo, con las técnicas de neuroimagen, como la tomografía por emisión de positrones (TEP), la resonancia magnética funcional (RMf) -que mejora a la anterior en que tienen una mayor resolución espacial y además no requiere de los trazadores radioactivos para su realización- y la magnetoencefalografía (MEG), técnicas todas el las no cruentas y nada o poco (caso de la TEP) invasivas.
Las estrategias de contrastación descritas no son excluyentes unas de otras. En la investigación psicobiológica se utilizan de forma combinada a la hora de investigar cualquier aspecto relacionado con el comportamiento humano, como puede ser la propia conducta sexual tratada anteriormente o, por ejemplo, los estudios sobre drogadicción en el ámbito de Psicofarmacología, bien en humanos o a través de modelos animales. Los equipos de investigación son interdisciplinares así como las técnicas utilizadas. No obstante, en las distintas disciplinas de la Psicobiología es posible encontrar preferencias en la utilización de unas u otras estrategias de investigación. Así, por ejemplo, la Psicofisiología utiliza la intervención conductual, la Neuropsicología, la Genética del Comportamiento, la Sociobiología y la Etología, la aproximación correlacional. Otras disciplinas psicobiológicas, tales como la Psicología Fisiológica, la Psicobiología del desarrollo o la Psicofarmacología, utilizan con más frecuencia la intervención somática.
En casi un siglo y medio, las Ciencias Biológicas, la Neurociencia, la Psicología científica, la Psicobiología, han cambiado radicalmente la idea que tenemos acerca de nosotros mismos. El cerebro, el órgano en el que se puede decir nos alojamos protegidos del mundo exterior por el duro cráneo que lo envuelve (y que sin embargo, no nos libra de nuestros miedos, fantasmas, ansiedades y muchos otros trastornos que en su interior se producen), nos está descubriendo poco a poco todos los circuitos y procesos que en su larga historia filogenética se han ido implementando en su interior para lograr una mejor representación del mundo exterior con el que tenemos que interactuar para satisfacer todas nuestras motivaciones, las que la filogenia ha creado para asegurar nuestra supervivencia y perpetuación, y las que nuestra vida y experiencia nos van generando al interaccionar con un entorno eminentemente social. Al final, todo el lo nos hace personas únicas, distintas a cualquier otra, que afrontamos la vida con diferente éxito según se hayan y hayamos combinado en nosotros todos los factores descritos. Esta es la savia que permea ya todas las disciplinas de la Psicología científica, aportando una perspectiva amplia y potente a cualquiera de los ámbitos en que se ejerza la práctica psicológica.
Cuando Mendel descubrió las leyes que llevan su nombre demostró que la verdadera herencia que cada progenitor trasmite a sus hijos es una dotación genética completa, un juego completo de genes -de los dos juegos que posee cada individuo en tanto que miembro de una especie diploide. Por su parte, Darwin cayó en la cuenta de que la conducta es un factor importante para la adaptación, medida por el éxito reproductivo; efectivamente, la conducta es variable y heredable, al menos hasta un cierto punto, como lo demuestra el éxito en la crianza selectiva de perros: es claro que la proliferación de razas de perros claramente diferenciadas tanto por su tamaño y aspecto como por su conducta tiene su explicación en la variabilidad genética existente en la especie Canis lupus; lo más probable es que la explicación de todas las diferencias entre perros y lobos sea de tipo comportamental, puesto que la diferencia esencial entre los perros y los demás cánidos tiene que ver con la capacidad de los perros para convivir pacífica y productivamente con los humanos (en esto consiste la domesticación), cosa que no cabe esperar de lobos, chacales, etc. Al parecer los híbridos de lobos y perros son fértiles, lo que apunta a que no se ha completado el proceso de especiación. Experiencias rusas en la crianza selectiva de zorros demuestra que es posible producir individuos manejables (que ni huyen ni atacan al hombre cuando éste se acerca) en un período corto de tiempo.
La teoría sintética de la evolución o neodarwinismo explica la evolución por selección natural como un cambio en las frecuencias alélicas de la población. Entonces, la cría selectiva que implicó la domesticación del lobo hasta convertirlo en perro, supuso un cambio en las frecuencias de determinados alelos, cambio genético que, supuestamente, explica las notables diferencias en la conducta de una y otra especie. Debemos aceptar, pues, que la conducta es un fenotipo bajo el cual subyace un genotipo que la explica (en mayor o menor medida dependiendo de cómo sea la interacción de esos genes con el ambiente). Igual que la diferencia de genotipos en los guisantes de Mendel explica las diferencias de color o textura de las semillas, una diferencia en los genotipos de lobos y perros debería explicar las diferencias entre ellos, tanto en apariencia física como en conducta. Lo realmente difícil en este caso, y en todo lo que tiene que ver con la explicación genética de la conducta, es definir el fenotipo: Mendel lo tuvo fácil con los guisantes, puesto que el color, la textura, etc. son rasgos discretos, fijos y fácilmente observables por cualquiera (objetivos), mientras que la conducta es algo continuo, variable y difícil de definir objetivamente -ambiguo en la medida en que cada observador podría dar una interpretación distinta del comportamiento observado.
No cabe duda de que hay una relación muy estrecha entre la genética molecular (estudio del material genético o ADN de los cromosomas y de los mecanismos bioquímicos que explican la expresión de la información contenida en ese ADN), genética mendeliana y teoría sintética de la evolución: efectivamente hoy por hoy sabemos de qué están hechos y cómo funcionan los genes cuyos efectos y modo de trasmisión Mendel describió, y cómo aquel los que favorecen el éxito reproductivo de sus portadores se implantan en la población mientras que los que proporcionan un rendimiento inferior en términos reproductivos tienden poco a poco a desparecer. Curiosamente, mucho antes de que los descubrimientos de Mendel fueran reconocidos por la sociedad, Galton (1822-1911 ), que, por cierto, era primo de Darwin, basándose en los principios de la teoría de la evolución del propio Darwin, dedujo que todos los rasgos conductuales humanos debían tener una base genética, resultado de la selección natural y, con este ligero bagaje teórico (ligero en la medida en que ni siquiera Darwin era capaz de explicar el origen material de las diferencias heredables, sobre las que, según él, actúa la selección natural), hipotetizó que la inteligencia humana tiene una base genética, y pretendió haberlo demostrado al comprobar que el grado de eminencia intelectual de los familiares masculinos de personajes eminentes era más probable cuanto mayor era el grado de parentesco familiar (biológico) de esos individuos eminentes con los familiares puestos a prueba: cuanto mayor era el grado de parentesco, mayor era la semejanza en eminencia intelectual. Por esto y por la enorme cantidad de investigaciones relacionadas que desarrolló o estimuló, se le considera a Galton el fundador de la Genética de la Conducta.
Estamos, pues, de acuerdo en que la conducta es un fenotipo, pero explicar la influencia de los genes sobre la conducta -el otro factor explicativo es, por supuesto, el ambiente- exige rastrear todo el proceso, que va desde el ADN de los gametos que se unen para formar el cigoto (el nuevo ser vivo) hasta el rasgo fenotípico conductual cuya base genética queremos demostrar. Para ello vamos a describir qué son los genes, cómo se heredan y cuales son los mecanismos bioquímicos que explican sus funciones.
Descubrimiento de la genética: Las Leyes de Mendel
Creo que no se puede ponderar el mérito de G. Mendel (1822-1884) como descubridor de las leyes de la herencia genética, como lo prueba la publicación en 1866 de su artículo «Experimentos en la hibridación de plantas»; y ello, a pesar de que tuvieron que pasar 35 años antes de que tales leyes llegaran a conocimiento del mundo. Y no deja de ser curioso que enmarque su investigación en el contexto de la fertilización artificial: no en vano hacía poco que se había demostrado la reproducción sexual de las plantas. No es que fuera desconocido el hecho de que los descendientes se parecen a sus progenitores y a veces incluso más a algún abuelo: el problema era que no había ninguna explicación material para dicha semejanza y además, parecía casi evidente que los descendientes presentan muy a menudo una apariencia intermedia de los rasgos de ambos progenitores, lo que parecía avalar la hipótesis de una herencia promedio. Pero Mendel se convenció de que, puesto que los caracteres (hoy diríamos rasgos) de una raza establecida (raza pura) se mantienen constantes sólo si tanto el polen como el óvulo proceden de plantas con dichos caracteres, ambos gametos debían aportar algo a los rasgos de la planta: a ese algo Mendel lo llamó factor (pero en la actualidad lo llamamos gen). Gracias a sus experimentos con polinización artificial cruzada entre variedades puras de guisantes (Pisum sativum) demostró que dichos factores no se mezclan, sino que se conservan íntegros como unidades de trasmisión hereditaria. Y ello a pesar de que «Los experimentos llevados a cabo en años anteriores con plantas ornamentales ya han proporcionado evidencia de que los híbridos, por regla general, no son exactamente un intermedio entre las especies progenitoras. Con algunos de los caracteres más notables, aquellos, por ejemplo, relacionados con la forma y tamaño de las hojas, la pubescencia de las diversas plantas, etc. el intermedio, en efecto, era casi siempre observado;» pero la clave de su trabajo estuvo en que «en otros casos, sin embargo, uno de los dos caracteres parentales es tan preponderante que es difícil o casi imposible, detectar el otro en el híbrido».
Si Mendel fue capaz de describir las leyes de la herencia genética fue gracias a una estrategia experimenta l modélica. Partió de la observación empírica de que cuando se cruzan en sucesivas generaciones individuos híbridos, hay rasgos que desaparecen y vuelven a aparecer: y puesto que «ningún (experimento) ha sido realizado hasta tal punto y de tal manera que permita determinar el número de formas diferentes bajo el cual la descendencia de los híbridos aparece, o para organizar estas formas con certeza de acuerdo con sus linajes separados, o definitiva mente para determinar sus relaciones estadísticas», Mendel aplicó la exhaustividad para describir matemáticamente qué pasa cuando se cruzan en sucesivas generaciones los híbridos originados a partir de dos variedades puras.
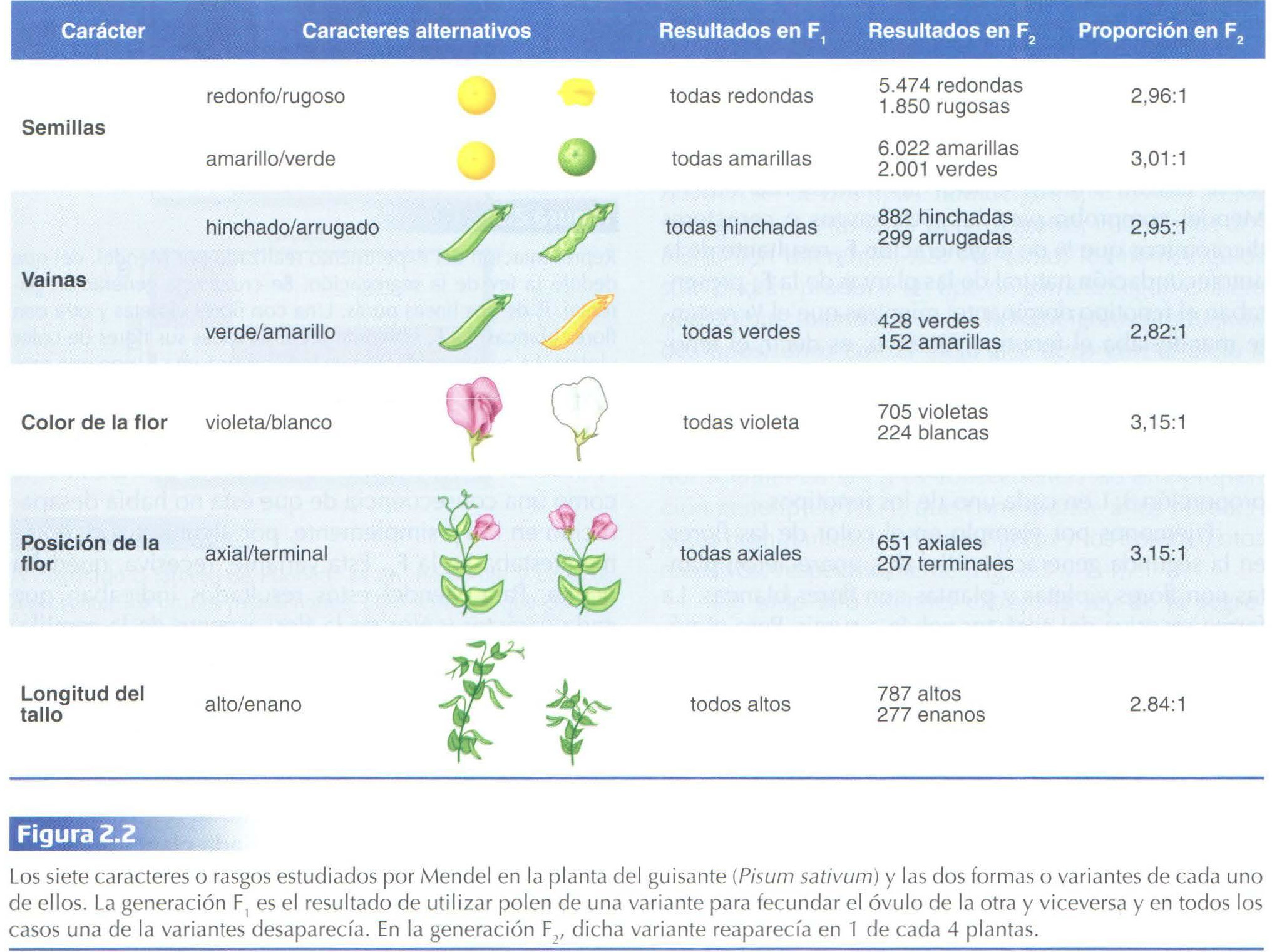
El paciente monje austríaco que fue G. Mendel, debe su fama (póstuma) y su éxito en el campo de lo que luego se conocería como Genética, no sólo a su meticulosidad experimental sino también a la toma de afortunadas decisiones a la hora de diseñar y analizar sus experimentos: su hipótesis le llevó a fijarse en rasgos discretos o cualitativos, para lo que eligió como material de estudio diversas variedades puras de plantas de guisantes en las que se podían discernir con claridad rasgos dicotómicos (Fig. 2.2) Que se trataba de variedades (razas) puras lo prueba el hecho de que durante 8 generaciones la autopolinización de cada raza dio siempre descendientes idénticos (fenotípicamente) a sus progenitores. De hecho estas razas puras le sirvieron de grupo control respecto a los grupos de plantas híbridas, que le permitieron demostrar las posteriormente llamadas Leyes de Mendel.
El primer paso consistió en cruzar entre sí dos razas puras que diferían en uno de los rasgos concretos (generación parental P), es decir, plantas de semilla lisa con plantas de semilla rugosa; plantas altas con plantas enanas, etc (hasta cubrir los siete rasgos que se enumeran en la Fig. 2.2 ): cruce monohíbrido. Así comprobó que todos los descendientes de estos cruces (generación F1) presentaban una apariencia en el rasgo considerado, idéntica a uno de los dos progenitores, independientemente además de si era el que aportó el óvulo o el polen. A la forma del carácter que se manifiesta (fenotipo) en esta generación F1 consideró Mendel calificarlo como dominante. Así pues, del cruce de plantas altas con plantas enanas resultó una planta híbrida donde el 100% fueron altas (tan altas al menos como las plantas de la raza alta utilizada en la fecundación ); y lo mismo ocurrió para los otros 6 rasgos dominantes (Fig. 2.2 ). A la variante del carácter que no aparece en esta generación F1 la llamó recesiva. Por tanto, a juicio de Mendel, para estos 7 caracteres, no se produce el efecto de mezcla hereditaria que cabía esperar.
Ley de la Segregación
El paso siguiente es comprobar qué ocurre cuando se cruzan entre sí al azar las plantas híbridas F1.
Mendel comprobó para los siete rasgos o caracteres dicotómicos que 3/4 de la generación F2 resultante de la autofecundación natural de las plantas de la F1 presentaban el fenotipo dominante, mientras que el 1/4 restante manifestaba el fenotipo recesivo, es decir, el fenotipo que no aparecía en la generación F1 (ver Fig. 2.3 para el caso del color de la flor). El mérito de Mendel estuvo en la interpretación matemática de estos datos cuant itativos, al establecer que la clave estaba en la proporción 3:1 en cada uno de los fenotipos.
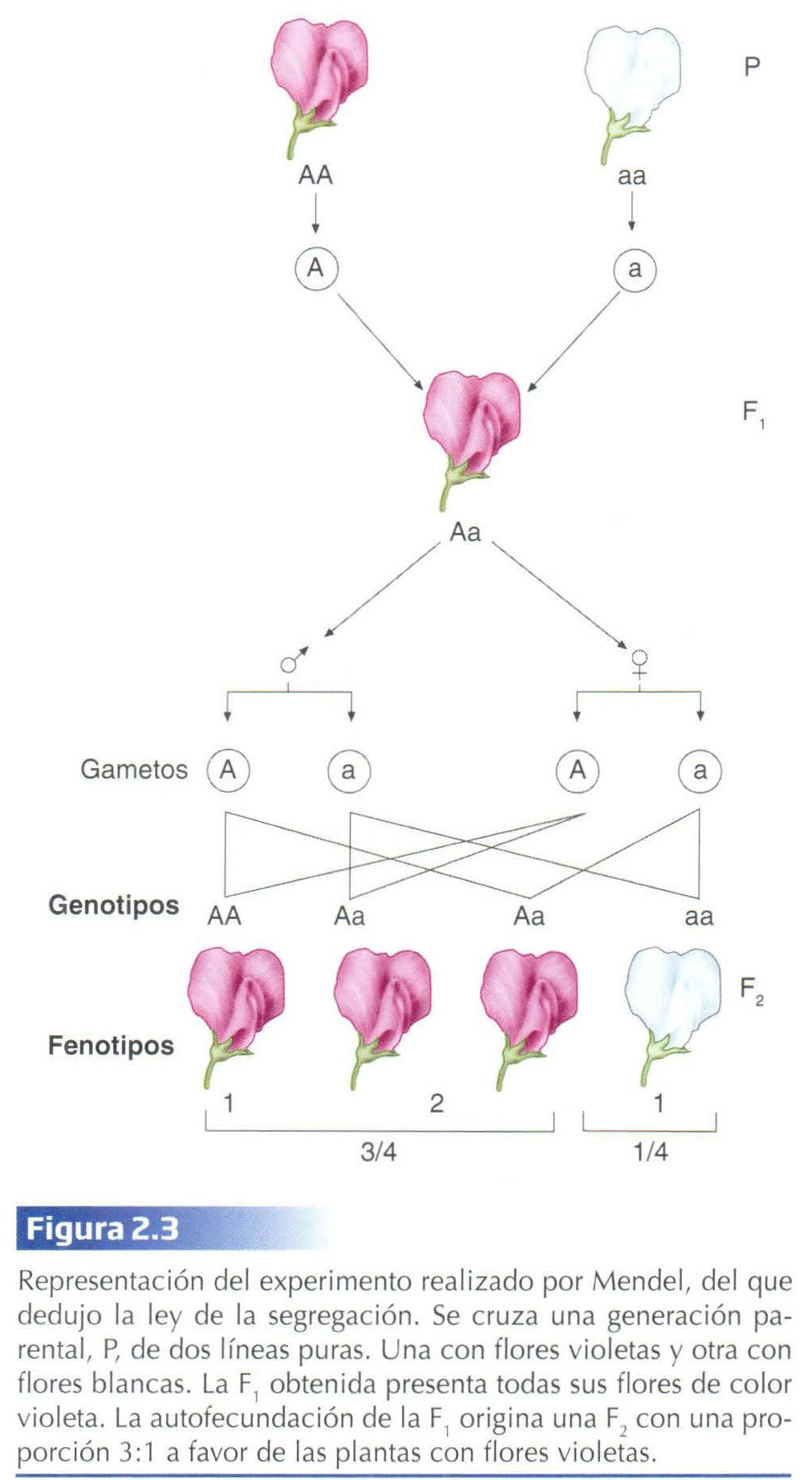
Fijémonos por ejemplo en el color de las flores: en la segunda generación filial (F2), aparecieron plantas con flores violetas y plantas con flores blancas. La forma recesiva del carácter volvía a surgir. Pero el número de plantas que obtuvo de uno y otro color de la flor no fue similar. Del total de descendientes que obtuvo, 705 presentaban la flor violeta y 224 la flor blanca. De esta forma obtuvo una proporción 3 a 1 (3:1) a favor de las plantas con flores violeta (Fig. 2.3).
El hecho de que la forma recesiva del carácter reapareciese en la F2, Mendel lo interpretó acertadamente como una consec uencia de que ésta no había desaparecido en la F1: simplemente, por alguna razón, no se manifestaba en la F1. Esta variante, recesiva, quedaba oculta. Para Mendel estos resultados indicaban que cada carácter (color de la flor, aspecto de la semilla, etc) era debido a un elemento o factor hereditario que corresponde al término moderno de gen. Por ejemplo, el gen para el carácter color de la flor, existe en dos formas o variantes, la responsable del color violeta y la causante de que la flor sea blanca. A estos genes que presentan más de una variante se les llama alelomorfos o, simplemente, alelos. Cada planta porta dos genes para cada carácter, uno procedente de la planta materna y otro de la paterna o, cuando hay autofecundación, del gameto femenino y del gameto masculino, respectivamente. En el caso que nos ocupa, el gen responsable del color de la flor es alelomorfo, presenta dos alelos, que podemos representar por la letra A, para el alelo dominante, y la letra a, para el recesivo. De esta forma, el alelo a es el responsable de que las flores presenten un color blanco y el alelo A de que lo presenten violeta. Las plantas con flores color violeta llevan dos alelos A (A A) o uno A y otro a (Aa) en el caso de los híbridos, mientras que las que presentan flores de color blanco llevarán dos alelos a (aa).
Mendel señaló que durante la formación de los gametos (gracias al proceso de meiosis que veremos enseguida) los alelos se separan (segregan), de tal forma que cada gameto recibe un solo alelo. Al juntarse dos gametos se restablece en el nuevo individuo la dotación doble habitual para cada carácter (Fig. 2.3 y 2.4).
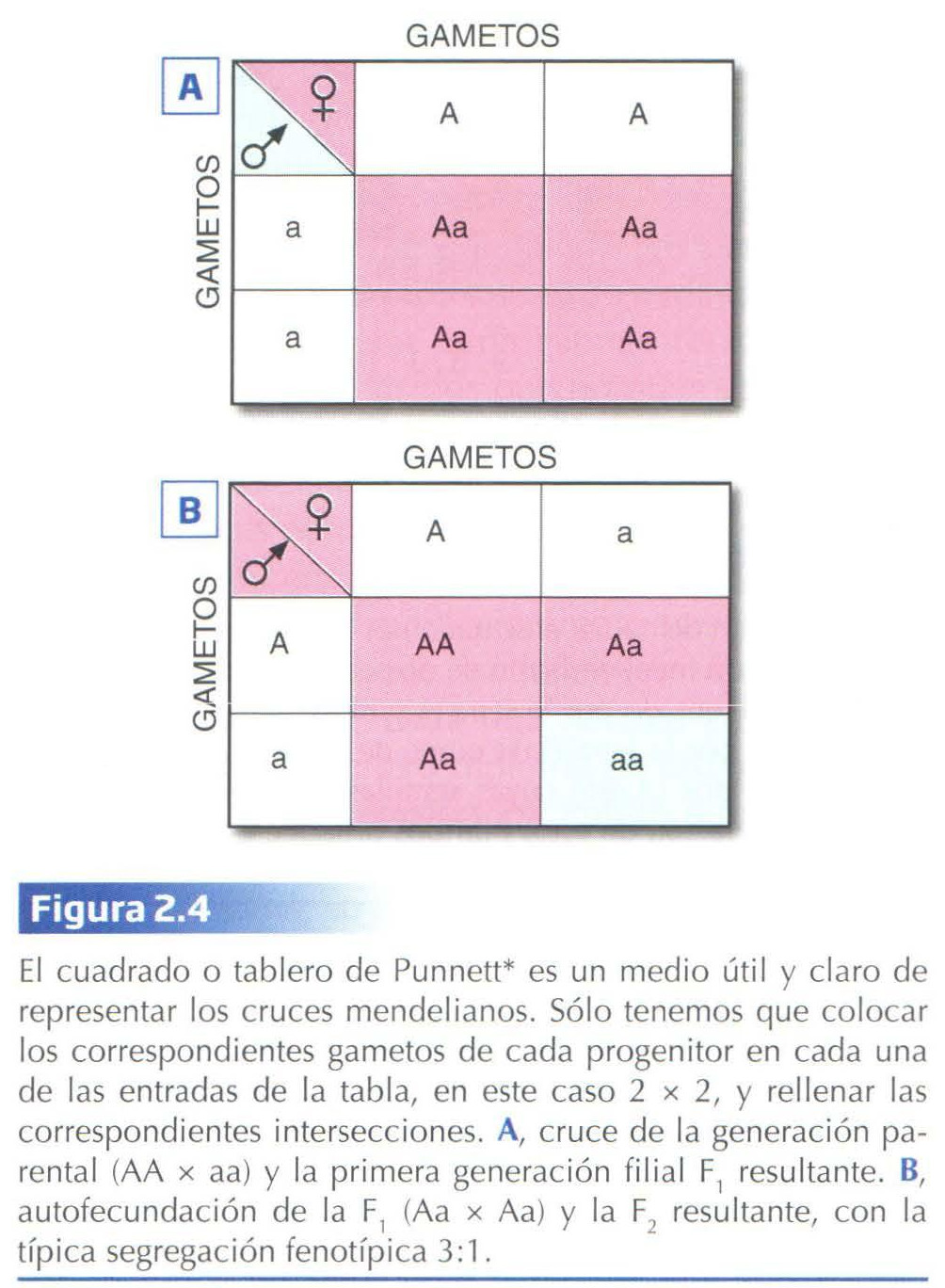
La constitución genética en relación a un carácter, o a todos los caracteres, se denomina genotipo y la manifestación externa del genotipo, fenotipo. Por ejemplo, el genotipo de un híbrido es Aa y su fenotipo, el color violeta (Fig. 2.3). Por su parte, los genotipos pueden ser de dos tipos: homocigotos, si los dos alelos son iguales, AA o aa, y heterocigotos, cuando los dos alelos son diferentes, Aa. Por tanto, los homocigotos sólo podrán producir un tipo de gameto según el alelo que portan, mientras que los heterocigotos producirán dos tipos, unos con el alelo A y otros con el alelo a (Fig. 2.4).
Teniendo en cuenta estos datos, se comprueba fáci lmente que la proporción fenotípica 3:1, obtenida por Mendel en la F2, es co nsecuencia de una proporción genotípica 1:2:1, que corresponde a los homocigotos dominantes, los heterocigotos y los homocigotos recesivos, respectivamente (Fig. 2.3 y 2.4).
De todo ello Mendel extrajo la ley de la segregación (primera ley de Mendel): las variantes recesivas enmascaradas en la F1 heterocigota, resultante del cruce entre dos líneas puras (homocigotas, por tanto), reaparecen en la segunda generación filial en una proporción 3:1, debido a que los miembros de la pareja de alelos del heterocigoto se separan sin experimentar alteración alguna durante la formación de los gametos.
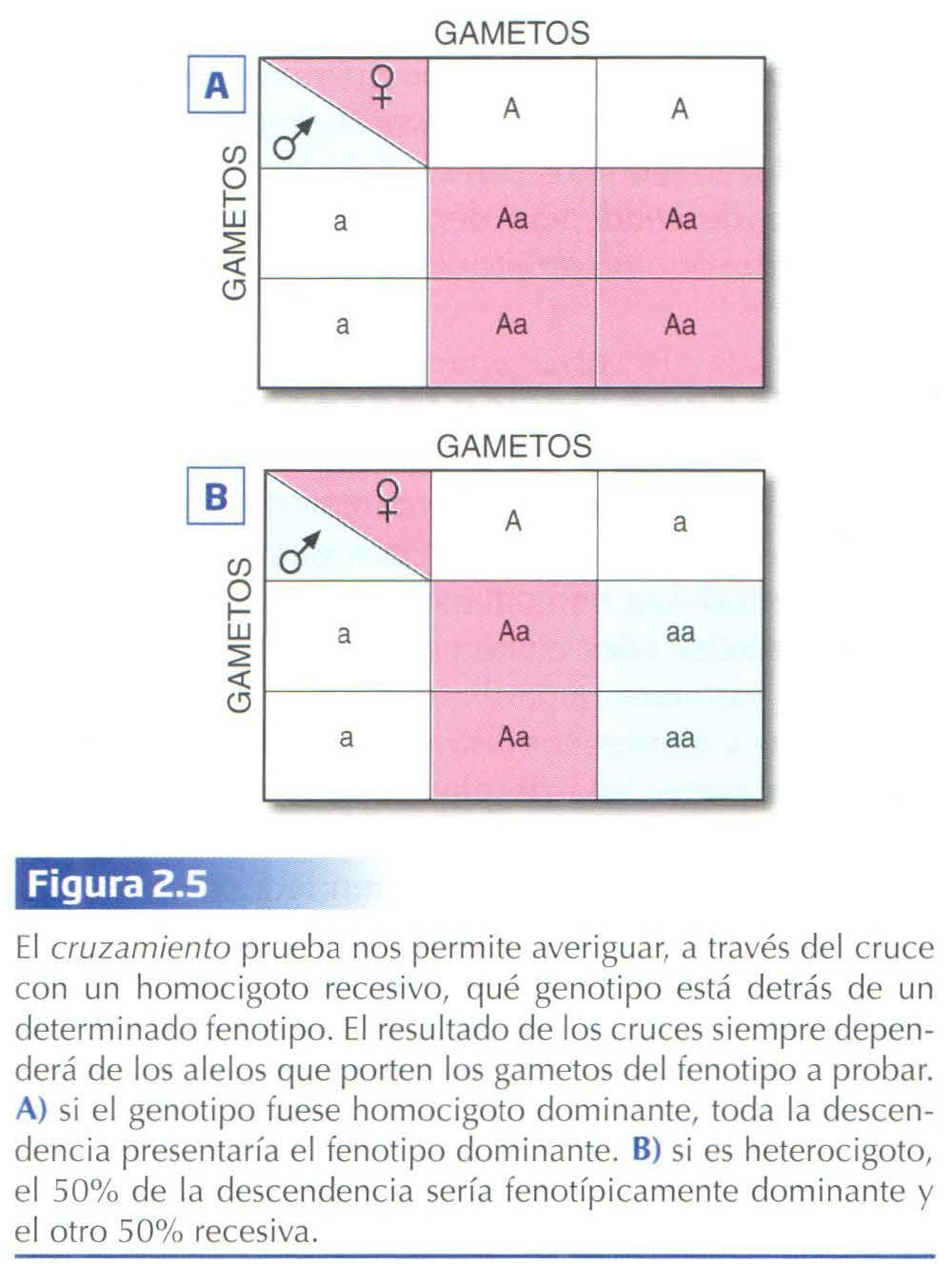
Dado que, fenotípicamente hablando, los homocigotos dominantes y los heterocigotos son indistinguibles, una manera de averiguar a qué genotipo corresponde un determinado fenotipo es a través del denominado cruzamiento prueba. Este consiste en cruzar individuos cuyo fenotipo queremos probar, con individuos homocigotos recesivos. Como éstos últimos sólo producen gametos con el alelo recesivo, el fenotipo de la descendencia dependerá únicamente del genotipo del otro progenitor (Fig. 2.5).
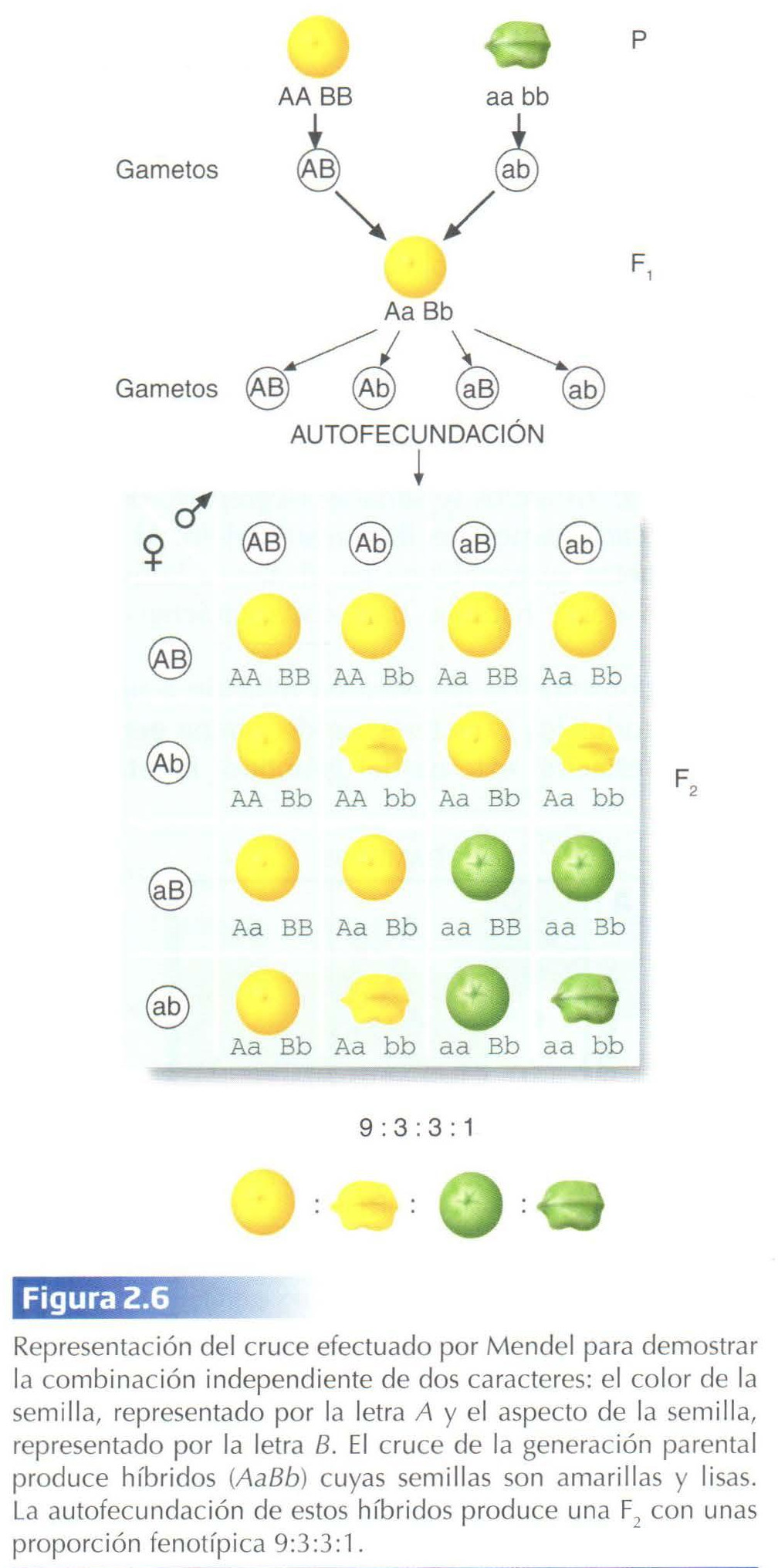
Ley de la Combinación Independiente
Una vez comprobadas las características de la herencia de un solo carácter con dos variantes o formas, Mendel estudió la herencia simultánea de dos caracteres diferentes, tales como el color de la semilla, que tiene dos variantes, amarilla o verde, y el aspecto de ésta, cuyas variantes son lisa y rugosa. Para ello cruzó dos líneas puras, una de plantas con semillas amarillas y lisas, y otra cuyas semillas eran verdes y rugosas (Fig. 2.6). Las plantas obtenidas en la F1 presentaban todas semillas amarillas y lisas. La primera ley seguía cumpliéndose para cada rasgo, ya que todos los individuos de la F 1 tenían el mismo fenotipo. Por otro lado, los resultados indicaban que tanto la variante amarilla como la lisa eran dominantes, mientras que la verde y la rugosa eran recesivas.
La autofecundación de las plantas de la F1 proporcionó una generación F2 constituida por las cuatro combinaciones posibles para los caracteres estudiados: semillas amarillas y lisas, amarillas y rugosas, verdes y lisas, y verdes y rugosas, con unas proporciones respectivas de 9:3:3:1 (Fig. 2.6). Considerados de forma independiente, cada carácter seguía presentándose en una proporción 3:1, es decir, se cumplía la ley de la segregación. Por otro lado, en la F2 habían aparecido combinaciones que no estaban presentes ni en la generación parental ni en la F1, es decir, plantas con semillas amarillas y rugosas y plantas con semillas verdes y lisas, lo cual implicaba que los caracteres color y aspecto de la semilla se habían transmitido de forma independiente. De estos resultados Mendel dedujo la ley de la combinación independiente: los miembros de parejas alélicas diferentes se segregan o combinan independientemente unos de otros cuando se forman los gametos.